Creier uman. Tesut nervos
Salut cititor ! Am ajuns în sfârșit... Ei bine, cum se poate face fără un site dedicat zonelor minții și lumii descriere detaliata ultimul? Pe de altă parte, nu aș vrea să transform paginile site-ului în paginile unui manual de anatomie a creierului. Este puțin probabil ca cei mai mulți să fie interesați de unde sunt localizați ventriculii, ce vine de unde și unde ies. Și, prin urmare, doar cele mai delicioase lucruri vor fi aici...
Funcțiile creierului:
Cap creier umanîndeplinește funcții mentale, de reglare și de coordonare. Este responsabil pentru tot ceea ce se întâmplă în organism, de la ciclu până la depunerea rezervelor de grăsime.
Răspunzător de munca coordonata toți mușchii corpului, de la mușchiul inimii până la mușchii sfincteri ai vaselor de sânge. De asemenea, responsabil pentru coordonarea mișcărilor și orientarea în spațiu.
Funcțiile mentale ale creierului constau în formarea unui fundal emoțional, semnale din aparatul senzorial și formarea unei imagini a lumii. De asemenea, creierul desfășoară procese de gândire, ia decizii, emite ipoteze și produce toate acestea sub forma unui discurs articulat, conștient.
Greutatea creierului
Greutatea creierului uman variază între 1,1 și 1,9 kg. Femeile au o greutate medie de 1,2 kg. , la bărbați în medie 1,4 kg. Dependenta abilități mentale bazat pe masa creierului nu a fost stabilit. Deși, cel care are mai multe șanse să devină un geniu creier mare(avizul experților). Majoritatea oamenilor de știință, politicieni, artiști și scriitori străluciți și remarcabili au avut un creier, o masă peste medie și, de asemenea, o arhitectură neobișnuit de complexă.
Structura creierului
Cuprinde creier mare, creier mic sau cerebel, sistem limbic (creier antic) și medular oblongata.
Creier mare
Sistemul limbic
Se compune din hipocamp, amigdala (amigdale), corpul calos, fornix, corpul mamilar, bulbul olfactiv, hipotalamus și talamus.
Sistemul limbic (creierul reptilian) este responsabil de comportamentul emoțional, comportamentul sexual, comportamentul agresiv - defensiv, motivația, învățarea și formarea memoriei. Participă la reglarea ciclului somn-veghe.
Următorul sistem fundamental este trunchiul cerebral.
Trunchiul cerebral
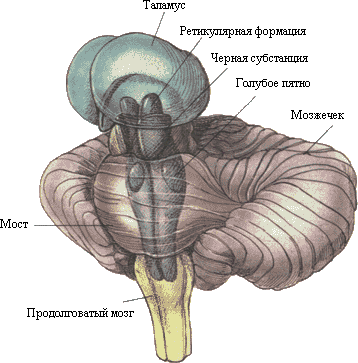 Cuprinde diencefal, mesenencefal, puț, medular oblongata și cerebel.
Cuprinde diencefal, mesenencefal, puț, medular oblongata și cerebel.
Responsabil cu organizarea reflexelor, implementarea diferite forme comportament, îndeplinește funcția conductivă, reglează ritmul cardiac, tensiune arteriala, ritmul respirator, reflexul de deglutitie.
Cerebel
Ne permite să facem mișcări, atât conștiente, cât și inconștiente, cu mare precizie. Ne permite să navigăm în spațiu și să menținem echilibrul.
Acesta este modul în care funcționează pe scurt creier uman. În general, se pare că am atins totul. De fapt, fiecare dintre departamentele și părțile creierului merită propriul articol. Creierul nostru conține o mulțime de lucruri interesante. Și, prin urmare, voi dedica o secțiune separată anatomiei creierului. În articolul următor voi vorbi despre. Deci nu pleca mult timp ;)
Creierul uman este cel mai complex mecanism biologic creat de natură. Are un potențial enorm, care probabil nu va fi niciodată pe deplin realizat. Viața misterioasă a materiei cenușii este uriașă Pata alba pe harta cunoaşterii umane. Cum funcționează creierul, cum funcționează - nici un singur locuitor al pământului nu poate da un răspuns complet și clar la aceste întrebări.
Totul despre creier este misterios: de la modul în care a apărut pe planeta albastră până la conexiunile sale cu lumea subtilă a Universului, care afectează direct profunzimile subconștientului unei persoane. Aceste ghicitori excită imaginația, îi îndeamnă pe oameni să caute altele noi și noi. metode neconvenționale studiu medular.
Se întâmplă că acest mecanism cel mai perfect este forțat să se studieze singur, dar procesul de cunoaștere, din păcate, nu are prea mult succes. Prea complexe, de neînțeles, diferite unele de altele și diverse sunt toate procesele care au loc în materie cenusie. Reflecțiile lor se regăsesc în lumea exterioară în fiecare secundă, oferind oamenilor posibilitatea de a trăi o viață interesantă, viață plină, să cunoască realitatea înconjurătoare și să-i admiri unitatea și lupta contrariilor.
ÎN corpul uman creierul ocupă o poziţie privilegiată. Din lumea de afarațesuturile sale cele mai delicate sunt protejate de craniu, în interior - lichidul cefalorahidian protejează în mod fiabil împotriva comoțiilor. Reprezentând doar două procente din greutate totală corpuri, acesta, pătat cu sute de mii vase de sânge organul absoarbe douăzeci la sută din oxigenul primit de plămânii noștri.
ÎN condiții extreme, când corpul moare de foame, creierul ia partea copleșitoare nutrienți. Dacă pierde cincizeci la sută din greutatea corporală, pierde doar cincisprezece la sută.
Pe partea de sus a creierului este acoperit cu un strat subțire gri, cu șanțuri și circumvoluții. Acesta se numește țesut nervos Cortex cerebral. Grosimea sa în diferite părți emisfere cerebrale variază de la 1,3 mm la 4,5 mm. Este format din paisprezece până la șaisprezece miliarde de neuroni, principalul element funcțional sistem nervos.
Aici centrul de gândire cu directă și părere, care se realizează prin mănunchiuri verticale de fibre. Informația vine de la simțuri la cortex prin impulsuri nervoase și semnale chimice. După procesare, este trimis înapoi sub formă de comenzi și servește drept ghid de acțiune pentru diferite părți ale corpului uman.
Cea mai mare parte a creierului (aproximativ 70%) este localizată în emisfere cerebrale. Ele sunt simetrice și conectate între ele prin corpul calos (un mănunchi de procese neuronale), care asigură schimbul de informații între ei.
Emisferele sunt formate din lobii frontal, temporal, parietal și occipital. ÎN Lobii frontali– centre care reglementează activitate motorie, V lobii parietali– zone de senzații corporale. Lobii temporali sunt responsabili de auz, centrii vorbirii, memorie, iar cei occipitali convertesc razele de lumină care lovesc retina în senzații vizuale.
Sub cortex se află nucleii creierului, care sunt formați din grupuri de neuroni, cum ar fi hipotalamusul și talamusul. Hipotalamus- o zonă mică a creierului care controlează funcțiile homeostatice ale corpului. talamus responsabil pentru starea de veghe și atenție.
Responsabil pentru poziția capului, a trunchiului și a membrelor, adică pentru a se asigura că o persoană se simte confortabil în timp ce stă vertical pe sol. cerebelul care se ascunde sub lobii occipitali emisfere cerebrale. De asemenea, joacă un rol decisiv în dezvoltarea diferitelor abilități necesare vieții de zi cu zi.
Greutate medie creierul adult un kilogram și jumătate. Există exemplare individuale de substanță cenușie care cântăresc două kilograme. Dar volumul și masa mare nu sunt în niciun caz semne ale unei minți extraordinare și ale unui intelect puternic. Există criterii complet diferite la lucru aici, care nu au fost încă studiate practic.
Creierul, în general, este un mecanism biologic foarte greu de studiat. Este prea complex și misterios să le dezvălui toate secretele pelerinilor în Țara Conștiinței exact așa.
Emisferele creierului
De exemplu, emisfera stângă și dreaptă– este ca două creiere într-un singur craniu. Fiecare dintre ei își gestionează propriile afaceri, dar în același timp își ajută colegul. Stânga se ocupă cu logica, gândire abstractă, dreapta – concret, figurat.
Dacă preia controlul asupra psihicului emisfera stângă, atunci starea de spirit a persoanei norocoase se va îmbunătăți. Va deveni prietenos, optimist, moale și vesel. Dar dacă dreapta începe să domine, atunci vâslele se vor usca. Depresia, iritabilitatea, izbucnirile de furie, agresivitatea sunt frecvente în acest caz.
De asemenea, este interesant faptul că specializarea emisferelor la bărbați este mult mai pronunțată decât la sexul frumos. Până la vârsta de șase ani, băieți emisfera dreaptă preia în totalitate funcţiile care îi sunt atribuite. Dar la fete rămâne mai plastic pentru o lungă perioadă de timp. Aproape de-a lungul vieții, femeile au capacitatea de a percepe spațial lumea înconjurătoare în in aceeasi masura comune ambelor jumătăți ale creierului.
O astfel de specificitate universală poate juca un rol pozitiv în Vătămarea corporală una dintre emisfere. A doua emisferă va prelua cu calm îndeplinirea funcțiilor pierdute ale fratelui său. Deci bărbații nu pot decât să invidieze.
Un mare interes pentru studierea activității creierului este acordat sentimentelor, gândurilor, emoțiilor, care în toată diversitatea lor enormă sunt caracteristice doar coroanei naturii, adică tu și eu. Animalele, deși au și materie cerebrală, nici măcar nu sunt aproape de oameni.
Viața spirituală este o consecință a muncii creierului, care sunt procese pur fizice și chimice sau altceva, misterioasă și de neînțeles? Această întrebare i-a îngrijorat mereu pe oameni, dar încă nu există un răspuns la ea.
În secolul al XIX-lea, rectorul Seminarului Teologic din Kiev, arhimandritul Boris, și-a subliniat părerile despre această problemă în eseul „Despre imposibilitatea unei explicații pur fiziologice a vieții mentale umane”. Ministru de rang înalt biserică ortodoxă, fiind de acord că viata mentala- aceasta este opera creierului, in acelasi timp el a sustinut ca fenomenele mentale isi au adevarata existenta in afara creierului. Atunci unde? „Acest lucru ne este necunoscut, deoarece este revelația lui Dumnezeu.”
De dragul obiectivității, trebuie spus că oamenii de știință sunt în mare măsură de acord cu slujitorul lui Dumnezeu. De exemplu, fiziologul englez C. Sherrington credea că gândirea se naște în afara materiei, dar apare în capul oamenilor, inducându-i astfel în eroare că ei înșiși au adus-o pe lume.
Dar anatomistul australian F. Hallem a încercat să explice acest mister din punct de vedere materialist. El a susținut că viața noastră spirituală este reprodusă în cortexul cerebral. Cu toate acestea, această formulare a întrebării nu a condus la nimic bun. Cufundat cu capul înainte în studiu procese fiziologice, încercând să atragă la ei bogățiile spirituale ale unei persoane, acest om învățat a mers până acolo încât a creat frenologia - o știință conform căreia se poate judeca caracterul oamenilor după configurația craniului. Ulterior, această ipoteză a fost adoptată de rasiști de toate dungile și nuanțe.
Creierul nu este sensibil la durere. Poate fi iritat soc electric, tăierea cu un bisturiu - o persoană nu o va simți. De ce o natură atât de rațională și practică nu s-a ocupat de cele mai importante functie de protectie pentru cel mai important organ al corpului nostru? Aparent pentru că materia cenușie nu poate fi restabilită. Odată ce este deteriorat, nimic nu poate fi reparat.
Dar fiecare nor are o căptușeală de argint. Lipsa efectului durerii a permis cercetătorilor materiei cenușii să folosească electricitatea în munca lor. Prin implantarea celor mai subțiri electrozi în diferite părți ale creierului, ei au putut afla cum funcționează părțile individuale ale acestuia și de ce sunt responsabili.
Dacă atingeți neuronii cu un electrod regiune temporală cortexul cerebral, atunci subiectul poate izbucni în amintiri (să zicem, din copilăria îndepărtată) care ar fi pur și simplu imposibile în condiții normale. Iritarea hipotalamusului va provoca agresivitate, iar dacă un electrod este implantat în formatiune reticulara, atunci poți gestiona frica.
Creierul tinde să-și amintească organele pierdute. O persoană își pierde un braț, după aceea trec ani, iar membrul amputat continuă să „trăiască” și să „doare insuportabil”. Astfel de dureri se numesc dureri fantomă și sunt bine cunoscute medicilor. Apropo, doar implantarea electrozilor vă permite să scăpați pentru totdeauna de acest factor neplăcut.
Așa, în schiță generală, cum funcționează creierul uman. În concluzie, aș dori să spun câteva cuvinte despre lucruri destul de ciudate care, deși foarte rar, sunt observate la indivizi individuali. Aceasta este absența materiei din creier. În timpul unei autopsii, în loc de neuroni și celule gliale, în craniul unei astfel de persoane se găsește apă obișnuită.
Astfel, patologul german Joachim Hoffmann, în timp ce autopsia cadavrul unui pacient care a suferit o tulburare psihică în timpul vieții, a descoperit o masă lichidă în capul său în locul imaginii obișnuite. Venerabilul doctor a fost șocat până la capăt, dar nu a putut explica acest fenomen.
Iată un alt exemplu. Un student englez care a venit acasă de sărbători a mers la spitalul local cu o plângere gravă durere de cap. Medicii nu au putut determina cauza mult timp stare proastă răbdător, dar după raze X creierele erau îngrozite. Acesta are tânăr materia cenușie a lipsit cu desăvârșire: în loc de ea, s-a stropit cu lichid. Este interesant că tânărul s-a comportat destul de adecvat și a fost în stare bună la universitate și a studiat destul de bine.
Nu este un secret pentru nimeni că, atunci când au deschis craniul „liderului proletariatului mondial”, domnul Ulyanov V.I (Lenin), nici luminari medicali ruși nu au găsit substanță cenușie în capul său. În loc de miliarde și miliarde de celule nervoase, în capul teroristului bolșevic era apă.
Creier uman– acesta este cel mai perfect, cel mai bine reglat mecanism biologic. Nu este nimic de prisos în ea, dar este necesar și chiar necesar oameni moderni Se utilizează doar 10%. Până la 90% din materia cenușie este nefolosită de-a lungul vieții. O cantitate mare neuronii nu sunt niciodată puși în funcțiune și nu beneficiază o persoană.
Care este acest beneficiu? Nu există un răspuns clar aici. Poate e intuiție genială, poate teleportare. Memoria ideală, perfecțiunea spirituală, cunoașterea universală nu pot fi excluse. Dacă toate acestea se află foarte aproape, sub craniu, atunci trebuie să lucrezi și să lucrezi pe tine pentru a-i trezi pe cei care dorm puteri extraordinare, care poate schimba radical viața fiecăruia dintre noi și, prin urmare, a întregii umanități în ansamblu.
Baza chimică a activității vitale a țesutului nervos are, pe de o parte, trăsături comune inerente celulelor oricărui țesut, iar pe de altă parte, caracteristici specifice determinate de natura funcțiilor îndeplinite de sistemul nervos în întregul organism. . Aceste caracteristici se manifestă atât în compoziția chimică, cât și în metabolismul țesutului nervos.
Tesut nervos este format din trei elemente celulare: neuroni (celule nervoase); neuroglia - sisteme de celule care înconjoară imediat celulele nervoase din creier și măduva spinării; elemente mezenchimale, inclusiv microglia (celule Gortega).
Cea mai mare parte a creierului este primele două tipuri de elemente celulare. Neuronii sunt concentrați în substanța cenușie (60-65% din materia creierului), în timp ce substanța albă a sistemului nervos central și nervi periferici constau în principal din elemente de neuroglia și derivatul lor - mielina.
STRUCTURA NEURONALE
Un neuron este format dintr-un corp celular, numeroase procese scurte ramificate - dendrite și unul proces lung- un axon, a cărui lungime poate atinge câteva zeci de centimetri (Fig. 134).
Volumul de citoplasmă conținut în procesele unei celule nervoase poate fi de câteva ori mai mare decât cantitatea sa din corpul celular. Corpul neuronului este înconjurat de o membrană plasmatică - plasmalemă (Fig. 135). În strânsă legătură cu plasmalema 1 din corpul neuronului și segmentele proximale ale dendritelor există așa-numita structură a membranei subterane. Acestea sunt rezervoare care sunt situate paralel cu suprafața membranei plasmatice și sunt separate de aceasta printr-o zonă de lumină foarte îngustă. ( 1 Când un neuron este excitat, permeabilitatea membranei plasmatice se modifică.)
Se presupune că tancurile se joacă rol importantîn metabolismul neuronilor. Ultrastructura principală a citoplasmei unui neuron este reticulul endoplasmatic (sau reticulul ergastoplasmatic). Reticulul endoplasmatic este un sistem de vezicule, tuburi și saci turtiți sau cisterne delimitate de membrană. Membranele reticulului endoplasmatic sunt conectate într-un anumit fel cu plasmalema și membrana nucleului neuronului.
Granulele localizate pe membranele reticulului endoplasmatic, precum și localizate liber în citoplasmă, sunt ribozomi.
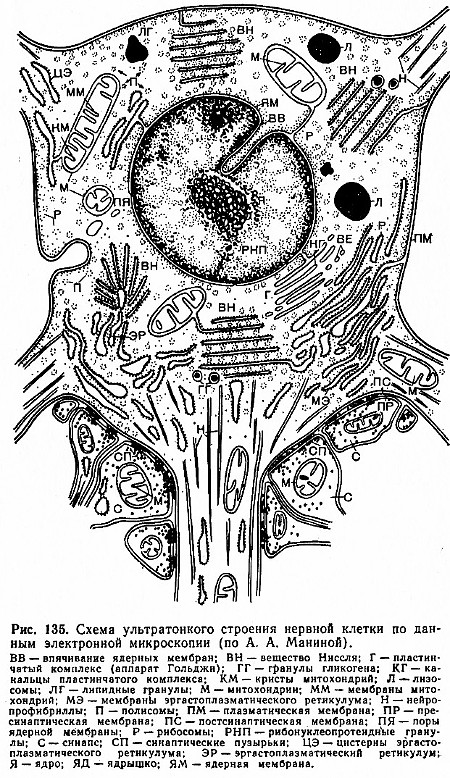
Unitatea structurală caracteristică a unei celule nervoase sunt corpurile Nissl, formate din acizi ribonucleici și proteine. Citoplasma dezvăluie, de asemenea, o rețea de filamente subțiri - neurofibrile, care împreună formează o rețea densă. Neurofibrilele sunt expresia structurală a orientării liniare corecte a moleculelor de proteine.
O componentă importantă a citoplasmei neuronului este complexul lamelar (aparatul Golgi), unde componentele lipidice ale celulei sunt concentrate în principal. Una dintre caracteristicile mitocondriilor izolate din celulele nervoase este că acestea conțin mai puține enzime implicate în procesele de oxidare. acizi grașiși aminoacizi decât mitocondriile din alte țesuturi.
În sistemul nervos central, lizozomii se găsesc în mod constant și îndeplinesc aceleași funcții ca lizozomii în alte organe și țesuturi.
Dimensiunea nucleului neuronului variază de la 3 la 18 microni, ajungând la 1/4 din dimensiunea corpului lor în neuronii mari.
STRUCTURA MIELINEI
Fibrele nervoase formate din axonii celulelor nervoase pot fi împărțite în două tipuri în funcție de structura lor: pulpoase (mielinizate) și fără pulpă (sărace în mielină).
Sistemul de conducere al sistemului nervos somatic, precum și sistemul nervos central, aparțin primului tip, care este funcțional mai avansat și are capacitatea de a transmite impulsuri nervoase cu viteză mare.
Substanța mielină este un concept pur morfologic. În esență, mielina este un sistem format prin stratificarea în mod repetat a membranelor celulelor neurogliale 2 în jurul proceselor nervoase (în periferie). trunchiuri nervoase neuroglia sunt reprezentate de lemocite, sau celule Schwann, iar în substanța albă a sistemului nervos central - astrocite macroglia). ( 2 Structură fină neuroglia sunt discutate în manuale speciale dedicate histologiei și morfologiei sistemului nervos.)
De compoziție chimică substanța mielină este un complex complex proteic-lipidic. Lipidele reprezintă până la 80% din reziduul dens; 90% din toate lipidele mielinei sunt reprezentate de colesterol, fosfolipide și cerebrozide.
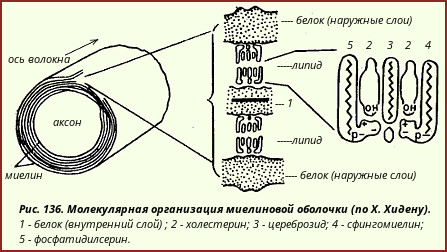
Există motive să credem că în straturile lipoide ale tecilor de mielină, moleculele diferitelor lipide au un aranjament strict definit (Fig. 136).
COMPOZIȚIA CHIMĂ A CREIERULUI
Substanța cenușie a creierului este reprezentată în principal de corpurile celulare ale neuronilor, iar substanța albă de axoni. În acest sens, aceste părți ale creierului diferă semnificativ în compoziția lor chimică. Aceste diferențe sunt în primul rând cantitative. Conținutul de apă din substanța cenușie a creierului este vizibil mai mare decât în substanța albă (Tabelul 54).
În substanța cenușie, proteinele reprezintă jumătate din substanțele dense, iar în substanța albă - o treime 3. Ponderea lipidelor în substanța albă reprezintă mai mult de jumătate din reziduul uscat, în substanța cenușie - doar aproximativ 30%. ( 3 Când sunt calculate pe baza greutății umede a țesutului, proteinele sunt distribuite aproximativ egal între materia cenușie (8%) și cea albă (9%) a creierului.)
Proteinele creierului
Proteinele reprezintă aproximativ 40% din masa uscată a creierului. Țesutul cerebral este un obiect dificil de studiat compoziția proteinelor din cauza conținut grozav lipide și prezența complexelor proteino-lipidic.
Pentru prima dată A. Ya Danilevsky a separat proteine țesut cerebral la solubil în apă şi soluții saline proteine si proteine insolubile. Cercetări ample în acest domeniu au fost efectuate și de către A. V. Palladin și colegii de muncă, care au împărțit proteinele țesutului nervos în patru fracții: extractibile cu apă, soluție KC1 4,5%, soluție NaOH 0,1% și reziduu insolubil. S-a stabilit că materia cenușie este mai bogată în proteine solubile în apă decât substanța albă: 30%, respectiv 19%. materie albă, dimpotrivă, conține mult mai mult (22%) reziduuri proteice insolubile decât materia cenușie (5%).
Ulterior, au fost izolate 5-10 fracții de proteine solubile ale creierului, diferind prin mobilitatea lor electroforetică.
În prezent, prin combinarea metodelor de extracție cu soluții tampon, cromatografie pe coloane DEAE-celuloză și electroforeza disc în gel de poliacrilamidă, a fost posibilă izolarea a aproximativ 100 de fracții proteice solubile diferite din țesutul cerebral.
Țesutul nervos conține atât proteine simple, cât și complexe. Proteine simple - albumine (neuroalbumine), globuline (neuroglobuline), proteine cationice (histone etc.) si proteine suport (neuroscleroproteine).
Întrucât albuminele și globulinele cerebrale sunt proprietati fizice si chimice oarecum diferite de proteinele serice similare, acestea sunt de obicei numite neuroalbumine și neuroglobuline. Cantitatea de neuroglobuline din creier este relativ mică - în medie 5% în raport cu toate proteinele solubile. Neuroalbuminele sunt componenta proteică principală a fosfoproteinelor din țesutul nervos, ele reprezintă cea mai mare parte a proteinelor solubile (89-90%). Neuroalbuminele sunt rare în stare liberă. Majoritatea neuroglobulinelor fac parte din proteine complexe. În special, se combină cu ușurință cu lipide, acizi nucleici, carbohidrați și alte componente non-proteice.
Proteinele care se deplasează la catod în timpul separării electroforetice la pH 10,5-12,0 sunt numite cationice. Principalii reprezentanți ai acestui grup de proteine în țesutul nervos sunt histonele, care sunt împărțite în cinci fracții principale în funcție de conținutul de reziduuri de lizină, arginină și glicină din lanțurile lor polipeptidice.
Neuroscleroproteine pot fi descrise ca proteine suport structural. Principalii reprezentanți ai acestor proteine sunt neurocolagenii, neuroelastinele, neurostrominele etc. Ele alcătuiesc aproximativ 8-10% din toate proteinele simple ale țesutului nervos și sunt localizate în principal în substanța albă a creierului și în sistemul nervos periferic.
Proteinele complexe ale țesutului nervos sunt reprezentate de nucleoproteine, lipoproteine, proteolipide, fosfoproteine, glicoproteine etc. Țesutul cerebral conține cantități semnificative de formațiuni supramoleculare și mai complexe, cum ar fi liponucleoproteine, lipoglicoproteine și, eventual, complexe lipogliconucleoproteice.
Nucleoproteine- proteine care aparțin fie dezoxiribonucleoproteinelor (DNPs), fie ribonucleoproteinelor (RNPs). Unele dintre aceste proteine sunt extrase din țesutul cerebral prin apă, o altă parte prin medii de sare și a treia cu 0,1 N. soluție alcalină.
Lipoproteinele conțin grupări lipidice. Ele alcătuiesc o parte semnificativă a proteinelor solubile în apă din țesutul cerebral. Componenta lor lipidica este formata in principal din fosfolipide si colesterol.
Proteolipide- singurele proteine complexe care pot fi extrase cu solvenți organici, de exemplu, un amestec de cloroform și metanol. Spre deosebire de lipoproteine, componenta lipidică din acestea predomină asupra componentului proteic. Cea mai mare cantitate proteolipidele sunt concentrate în mielină în cantități mici fac parte din membranele sinaptice și veziculele sinaptice.
Fosfoproteine- proteine complexe, a căror grupare protetică este adesea o grupare fosfat conectată printr-o legătură esterică la un rest serină. Conținutul de fosfoproteine din creier este mai mare decât în alte organe și țesuturi - aproximativ 2% în raport cu toate proteinele complexe ale creierului. Fosfoproteinele se găsesc în membranele diferitelor structuri morfologice ale țesutului nervos.
Glicoproteine reprezintă un grup extrem de eterogen de proteine. Pe baza cantității de proteine și carbohidrați care alcătuiesc glicoproteinele, acestea pot fi împărțite în două grupe principale. Primul grup este glicoproteinele, care conțin de la 5 până la 40% carbohidrați și derivații acestora; partea proteică este formată în principal din albumine și globuline. Glicoproteinele care alcătuiesc al doilea grup conțin 40-85%. carbohidrați, o componentă lipidă este adesea detectată; prin compoziţia lor pot fi clasificate ca glicolipoproteine.
| ÎN anul trecut Un număr de proteine specifice au fost găsite în țesutul nervos. Aceste proteine includ, în special, proteina S-100 și proteina 14-3-2. Proteina S-100, sau proteina lui Moore, se mai numește și proteină acidă deoarece conține un numar mare de reziduuri de acizi glutamic și aspartic. Această proteină este concentrată în principal în neuroglia (85-90%), în neuroni nu este mai mult de 10-15% din cantitatea totală din creier. S-a stabilit că concentrația proteinei S-100 crește în timpul dresajului (dresajului) animalelor. Cu toate acestea, nu există niciun motiv să credem că proteina S-100 este direct implicată în formarea și stocarea memoriei. Este posibil ca participarea sa la aceste procese să fie indirectă. Proteina 14-3-2 este, de asemenea, o proteină acidă. Spre deosebire de proteina S-100, este localizată în principal în neuroni; în celulele neurogliale conţinutul său este scăzut. Rolul proteinei 14-3-2 în îndeplinirea funcțiilor specifice ale țesutului nervos este încă neclar. |
Enzime.Țesutul cerebral conține un număr mare de enzime care catalizează metabolismul carbohidraților, lipidelor și proteinelor. Cu toate acestea, până acum doar două enzime au fost izolate sub formă cristalină din sistemul nervos central al mamiferelor - acetilcolinesteraza și creatinkinaza.
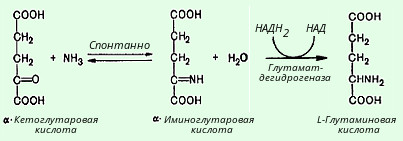
Un număr semnificativ de enzime în țesutul cerebral se găsesc în mai multe forme moleculare (izoenzime): lactat dehidrogenază, aldolază, creatin kinază, hexokinază, malat dehidrogenază, glutamat dehidrogenază, colinesterază, fosfatază acidă, monoaminoxidază și altele.
| Tabelul 55. Compoziția lipidică a țesutului nervos (după Norton, Poduslo, Suzuki, 1966) | |||
| materie cenusie | materie albă | Mielina | |
| Conținutul total de lipide, % din greutatea uscată | 32,7 | 54,9 | 70 |
| Ca procent din lipidele totale | |||
| Colesterolul | 22,0 | 27,5 | 27,7 |
| Cerebrozide | 5,4 | 19,8 | 22,7 |
| Gangliozide | 1,7 | 5,4 | 3,8 |
| Fosfatidiletanolamine | 22,7 | 14,9 | 15,6 |
| Fosfatidilcoline | 26,7 | 12,8 | 11,2 |
| Fosfatidilserine | 8,7 | 7,9 | 4,8 |
| Fosfatidilinozitoli | 2,7 | 0,9 | 0,6 |
| Plasmalogeni | 8,8 | 11,2 | 12,3 |
| Sfingomieline | 6,9 | 7,7 | 7,9 |
Lipidele
Printre componentele chimice ale creierului, un loc special îl ocupă lipidele, al căror conținut ridicat și natură specifică conferă țesutului cerebral. caracteristici. Grupul de lipide cerebrale include fosfolipide, colesterol, sfingomieline, cerebrozide, gangliozide și o cantitate foarte mică de grăsime neutră (Tabelul 55). În plus, multe lipide ale țesutului nervos sunt în strânsă relație cu proteinele, formând, în special, sisteme complexe precum proteolipidele.
În substanța cenușie a creierului, fosfolipidele reprezintă mai mult de 60% din totalul lipidelor, iar în substanța albă - aproximativ 40%. Dimpotrivă, substanța albă conține mai mult colesterol, sfingomieline și mai ales cerebrozide decât substanța cenușie.
Carbohidrați
Țesutul creierului conține glicogen și glucoză. Cu toate acestea, în comparație cu alte țesuturi, țesutul creierului este sărac în carbohidrați. Conținutul total de glucoză din creierul diferitelor animale este în medie de 1-4 µmol per 1 g de țesut, iar glicogenul - 2,5-4,5 µmol per 1 g de țesut (calculat ca glucoză). Este interesant de observat că conținutul total de glicogen din creierul embrionilor și al animalelor nou-născute este semnificativ mai mare decât în creierul adulților. De exemplu, la șoarecii nou-născuți, spre deosebire de adulți, nivelurile de glicogen sunt de 3 ori mai mari. Pe măsură ce creierul crește și se diferențiază, concentrația de glicogen scade rapid și rămâne relativ constantă la animalul adult.
Țesutul cerebral conține, de asemenea, produși intermediari ai metabolismului carbohidraților: hexoză și trioză fosfați, acizi lactici, piruvici și alți acizi. În tabel 56 prezintă date despre conținutul unor componente intermediare ale metabolismului carbohidraților în creierul șobolanilor.
| Tabelul 56. Date medii privind conținutul unor metaboliți ai metabolismului carbohidraților în creierul șobolanilor (conform Bergmeyer, 1970) | |
| metabolit | Conținut, µmol per 1 g greutate umedă de țesut |
| Glucoză-6-fosfat | 0,039-0,049 |
| Fructoza 6-fosfat | 0,017-0,023 |
| Fructoză 1,6-bifosfat | 0,010-0,017 |
| Fosfodioxiacetona | 0,024 |
| 3-fosfogliceraldehida | 0,021-0,046 |
| 3-fosfoglicerat | 0,085-0,100 |
| 2-fosfoglicerat | 0,010-0,016 |
| Fosfoenolpiruvat | 0,035-0,097 |
| piruvat | 0,120-0,190 |
| lactat | 1,26-1,70 |
Nucleotide de adenină și fosfat de creatină
Dintre nucleotidele libere din țesutul cerebral, nucleotidele de adenină reprezintă aproximativ 84%. Majoritatea nucleotidelor rămase sunt derivați de guanină. În general, numărul compușilor de mare energie din țesutul nervos este mic. Astfel, în creierul șobolanilor, conținutul mediu de nucleotide și creatină fosfat este (în µmol per 1 g greutate umedă): ATP-2,30-2,90; ADP-0,30-0,50; AMP-0,03-0,05; GTP-0,20-0,30; GDF-0,15-0,20; UTF-0,17-0,25; creatina fosfat - 3,50-4,75. Distribuția principalilor compuși macroergici este aproximativ aceeași în toate părțile creierului.
Conținutul de nucleotide ciclice (cAMP și cGMP) din creier este mult mai mare decât în multe alte țesuturi. Nivelul cAMP din creier este în medie de 1-2, iar cGMP până la 0,2 nmol per 1 g de țesut. Creierul este, de asemenea, caracterizat de activitate ridicată enzime ale metabolismului ciclic al nucleotidelor. Majoritatea cercetătorilor cred că nucleotidele ciclice sunt implicate în transmiterea sinaptică.
Minerale
Na, K, Cu, Fe, Ca, Mg și Mn sunt distribuite relativ uniform în creier între substanța cenușie și cea albă. Conținutul de fosfor în substanța albă este mai mare decât în substanța cenușie.
În tabel 57 prezintă date medii privind conținutul principalelor componente minerale din creier și fluide corporale.
După cum se poate observa din datele prezentate în tabel. 59, concentrația ionilor de potasiu, sodiu și clor din creier diferă mult de concentrația lor în fluidele corpului.
Raportul cantitativ dintre anionii anorganici și cationii din țesutul cerebral indică deficitul de anioni. Calculele arată că pentru a acoperi deficiența de anioni, ar fi nevoie de de 2 ori mai multe proteine decât sunt prezente în țesutul cerebral. În general, este acceptat că deficitul de anioni rămase este acoperit de lipide. Este foarte posibil ca participarea lipidelor la echilibrul ionic să fie una dintre funcțiile lipidelor în activitatea creierului.
CARACTERISTICI ALE METABOLISMULUI ȚESUTULUI NERVOS
BAZELE CHIMICE ALE APARIȚII ȘI CONDUITĂRII IMPULSULUI NERVOS
Care sunt bazele chimice pentru apariția și menținerea potențialelor bioelectrice (potențialele de repaus și de acțiune)? Majoritatea cercetătorilor sunt de părere că fenomenele de polarizare electrică a celulei sunt cauzate de distribuția neuniformă a ionilor de potasiu și sodiu pe ambele părți. membrana celulara. Membrana are permeabilitate selectivă: mai mare pentru ionii de potasiu și mult mai puțin pentru ionii de sodiu. În plus, există un mecanism în celulele nervoase care menține conținutul de sodiu intracelular la un nivel scăzut în ciuda gradientului de concentrație. Acest mecanism se numește „pompa de sodiu”.
În anumite condiții, permeabilitatea membranei la ionii de sodiu crește brusc.
La repaus partea interioară Membrana celulară este încărcată electronegativ în raport cu suprafața exterioară. Acest lucru se explică prin faptul că cantitatea de ioni de sodiu pompați din celulă folosind „pompa de sodiu” nu este exact echilibrată de aportul de ioni de potasiu în celulă. În acest sens, unii dintre cationii de sodiu sunt reținuți de stratul interior de contraioni (anioni) de pe suprafața exterioară a membranei celulare.
Când este excitată de unul sau altul agent, permeabilitatea membranei celulei nervoase (axon) se modifică selectiv: crește selectiv pentru ionii de sodiu (de aproximativ 500 de ori) și rămâne neschimbată pentru ionii de potasiu. Ca rezultat, ionii de sodiu se repetă în celulă. Fluxul compensator al ionilor de potasiu spre exterior din celulă este oarecum întârziat. Acest lucru are ca rezultat o sarcină negativă pe suprafața exterioară a membranei celulare. Suprafața interioară a membranei capătă o sarcină pozitivă; membrana celulară (în special, membrana axonală, adică fibra nervoasă) este reîncărcată și apare un potențial de acțiune, sau un vârf. Durata vârfului nu depășește 1 ms. Are o fază de creștere, o fază de vârf și o fază de descreștere. Faza descendentă (scădere potențială) este asociată cu o predominanță crescândă a producției de ioni de potasiu față de aportul de ioni de sodiu - potențial de membrană revine la normal. După ce impulsul este efectuat, starea de repaus este restabilită în celulă. În această perioadă, ionii de sodiu care au intrat în neuron în timpul excitației sunt înlocuiți cu ioni de potasiu. Această tranziție are loc împotriva gradientului de concentrație, deoarece există mult mai mulți ioni de sodiu în mediul extern din jurul neuronilor decât în celulă după momentul excitării acesteia. Tranziția ionilor de sodiu față de gradientul de concentrație, așa cum sa menționat deja, se realizează folosind o pompă de sodiu, a cărei funcționare necesită energie ATP. În cele din urmă, toate acestea duc la restabilirea concentrației inițiale de cationi de potasiu și sodiu în interiorul celulei (axon), iar nervul este gata să primească următorul impuls de excitare.
Un alt proces la fel de important pentru tesutul nervos este transmiterea impuls nervos de la o celulă nervoasă la alta sau un efect asupra celulelor unui organ efector.
Rolul mediatorilor în transmiterea impulsurilor nervoase
Creierul are miliarde de neuroni care comunică între ei prin neurotransmițători.
O substanță chimică poate fi clasificată ca mediator numai dacă îndeplinește o serie de criterii. Fibrele nervoase trebuie să conțină enzimele necesare sintezei acestei substanțe. Când nervii sunt iritați, această substanță trebuie să fie eliberată, să reacționeze cu un receptor specific de pe celula postsinaptică și să provoace reactie biologica. Trebuie să existe mecanisme care să oprească rapid efectele acestei substanțe chimice.
Toate aceste criterii sunt îndeplinite de două substanțe - acetilcolina și norepinefrina. Nervii care le contin se numesc colinergici, respectiv adrenergici. În conformitate cu aceasta, toate sistemele eferente sunt împărțite în colinoreactive și adrenoreactive.
O serie de altele substanțe chimice satisface multe, dar nu toate criteriile enumerate. Astfel de mediatori includ dopamina, adrenalina, serotonina, octopamina, histamina, acidul y-aminobutiric etc.
Grupul mare de sisteme colinoreactive este foarte eterogen atât structural, cât și funcțional. Aceste sisteme sunt unite de un mediator - acetilcolina - și de structura generală a sinapselor.
Acetilcolina este un ester al acidului acetic și al colinei. Se sintetizează în celula nervoasa din colină și forma activă de acetat - acetil coenzima A folosind o enzimă specială colin acetiltransferaza (colina acetilază):
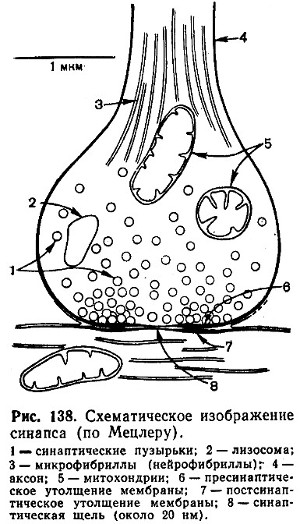
O sinapsă poate fi imaginată ca un spațiu îngust - un gol, limitat pe o parte de membrana presinaptică, pe de altă parte - de membrana postsinaptică (Fig. 138). Membrana presinaptică este formată dintr-un strat interior aparținând citoplasmei terminației nervoase și un strat exterior format din neuroglia. Membrana pe alocuri este îngroșată și compactată, în altele este subțiată și are găuri prin care citoplasma axonului poate comunica cu spațiul sinaptic. Membrana postsinaptică este mai puțin densă și nu are găuri. Sinapsele neuromusculare sunt construite într-un mod similar, dar au o structură mai complexă a complexului membranar.
În termeni generali, imaginea participării acetilcolinei la transmiterea excitației nervoase poate fi prezentată după cum urmează. În sinaptică terminații nervoase sunt bule (vezicule) cu diametrul de 30-80 nm care contin neurotransmitatori. Aceste vezicule sunt acoperite cu o înveliș, care este formată de proteina clatrină (gr. mol. relativă 180.000). La sinapsele colinergice, fiecare veziculă cu un diametru de 80 nm conține ~40.000 de molecule de acetilcolină. Când este excitat, mediatorul este eliberat „quanta”, adică prin golirea completă a fiecărei bule individuale. ÎN conditii normale sub influența unui impuls puternic, se eliberează aproximativ 100-200 de cuante ale transmițătorului - o cantitate suficientă pentru a iniția un potențial de acțiune în neuronul postsinaptic. Acest lucru se întâmplă aparent astfel: depolarizarea membranei terminalelor sinaptice determină un flux rapid de ioni de calciu în celulă. O creștere temporară a concentrației intracelulare a ionilor de calciu stimulează fuziunea membranei veziculelor sinaptice cu membrana plasmatică și astfel declanșează procesul de eliberare a conținutului acestora. Este nevoie de aproximativ patru ioni de calciu pentru a elibera conținutul unei vezicule. Acetilcolina eliberată în fanta sinaptică interacționează cu proteina chemoreceptor care face parte din membrana postsinaptică. Ca urmare, permeabilitatea membranei se modifică - debitul său pentru ionii de sodiu crește brusc. Interacțiunea dintre receptor și transmițător declanșează o serie de reacții care fac ca celula nervoasă postsinaptică sau celula efectoră să își îndeplinească funcția specifică. După eliberarea emițătorului, trebuie să înceapă o fază de inactivare sau îndepărtare rapidă a acestuia pentru a pregăti sinapsa pentru perceperea unui nou impuls. La sinapsele colinergice acest lucru se întâmplă în două moduri. În primul rând, acetilcolina suferă hidroliză enzimatică. A doua cale este transportul activ dependent de energie al acetilcolinei în neuron, unde se acumulează pentru reutilizare ulterioară.
Descompunerea hidrolitică a acetilcolinei în acid acetic iar colina este catalizată de o enzimă numită acetilcolinesterază:
| În majoritatea părților creierului, acetilcolina este hidrolizată de acetilcolinesteraza (o adevărată colinesterază care hidrolizează acetilcolina mai repede decât alți esteri de colină). Există și alte esteraze în țesutul nervos care sunt capabile să hidrolizeze acetilcolina, dar mult mai lent decât, de exemplu, butirilcolina. Aceste esteraze se numesc colinesteraza (sau pseudocolinesteraza). Sistemele colinergice includ neuronii motori care formează joncțiunea neuromusculară, toți neuronii preganglionari ai sistemului nervos autonom și neuronii postganglionari ai sistemului nervos parasimpatic. Un număr mare de zone simpatice colinergice se găsesc și în creier. În funcție de sensibilitatea la un anumit grup de compuși chimici, neuronii colinergici sunt împărțiți în „muscarinici” (activați de muscarină) și „nicotinici” (activați de nicotină). Receptorii muscarinici de acetilcolină găsiți în mulți neuroni sistem autonom, sunt blocate în mod specific de atropină. Sinapsele nicotinice sunt prezente în ganglioni și muschii scheletici. Inhibitorii lor sunt curarul și componenta activă a acestei otrăviri - D-tubocurarina. |
Trebuie subliniat că în sistemul adrenoreactiv există două tipuri de receptori pentru norepinefrină: receptori α- și β-adrenergici. Acești receptori se pot distinge unul de altul prin reacțiile specifice pe care le provoacă, precum și prin agenții specifici care pot bloca aceste reacții.
Receptorii β-adrenergici activează celula eferentă cu ajutorul adenozinei-3",5"-monofosfat sau cAMP - un "al doilea mesager" universal între hormoni și diverse funcții celule care sunt afectate de hormoni.
S-a stabilit că, de îndată ce receptorul β-adrenergic (situat pe suprafața exterioară a membranei celulare efectoare) începe să interacționeze cu norepinefrina, suprafata interioara membrana celulară, se activează enzima adenilat ciclază. Adenilat ciclaza apoi transformă ATP în cAMP în celulă; acesta din urmă, la rândul său, este capabil să influențeze metabolismul celular. Această serie complexă de reacții secvențiale poate fi blocată de propranolol, o substanță care interferează cu legarea norepinefrinei de receptorul β-adrenergic. Există o opinie că receptorul β-adrenergic nu este altceva decât subunitatea receptorului adenilat-ciclazei.
Se știe că enzima monoaminoxidaza (MAO) joacă un rol deosebit în metabolismul mediatorilor catecolaminei. Această enzimă elimină gruparea amino (-NH 2) din norepinefrină, serotonină, dopamină și adrenalină, inactivând astfel mediatorii menționați. Cu toate acestea, în ultimii ani s-a demonstrat că, pe lângă transformarea enzimatică, există un alt mecanism de inactivare rapidă, sau mai degrabă îndepărtarea, a mediatorilor. S-a dovedit că norepinefrina dispare rapid din fanta sinaptică ca urmare a absorbției secundare de către nervii simpatici; Din nou, în fibra nervoasă, mediatorul nu poate afecta în mod natural celulele postsinaptice. Mecanismul specific al acestui fenomen nu este încă pe deplin clar.
Sistemele adrenergice și colinergice ale creierului interacționează strâns cu alte sisteme ale creierului, în special cu sistemul serotonic. Neuronii care conțin serotonină sunt concentrați în principal în nucleele trunchiului cerebral. Rolul de neurotransmițător al serotoninei apare ca urmare a interacțiunii serotoninei cu receptorii serotoninergici specifici.
Studiile efectuate cu inhibitorul sintezei serotoninei p-clorofenilalanina, precum și cu alți inhibitori, sugerează că serotonina afectează procesele de somn. De asemenea, a fost dezvăluit că inhibarea activității secretoare a glandei pituitare de către corticosteroizi este mai puțin eficientă la acele animale al căror creier este mai sărac în serotonină.
Un neurotransmițător important care îndeplinește funcții inhibitorii este acidul γ-aminobutiric (GABA). Cantitatea de GABA din creier este de multe ori mai mare decât alți neurotransmițători. Astfel, în hipotalamus, conținutul total de acetilcolină, norepinefrină, dopamină și serotonină nu depășește 10 mcg/g, în timp ce GABA în această parte a creierului este mai mare de 600 mcg/g.
În prezent, un număr mare de medicamente, care actioneaza printr-un sistem de mediatori. Mulți substanțe medicinale, utilizate cu succes în tratamentul hipertensiunii arteriale, afectează acumularea și eliberarea de mediatori adrenergici. De exemplu, rezerpina este o scădere presiunea arterială medicamentul inhibă în mod specific procesul de transfer al catecolaminelor către granule speciale de neuroni și, prin urmare, face aceste amine disponibile pentru acțiunea MAO endogenă.
Hipotensiv medicamentele, precum α-metildopa, sub influența enzimelor conținute în celula nervoasă (axon) sunt transformate în substanțe asemănătoare norepinefrinei în structura lor. Acești neurotransmițători „falși” se acumulează și sunt eliberați împreună cu neurotransmițătorii naturali, diluându-i și reducându-le astfel efectul.
Multe antidepresive (substanțe care ameliorează depresia) cresc conținutul de catecolamine în fanta sinaptică, adică crește cantitatea de mediator pentru stimularea receptorului. Astfel de substanțe, în special, includ imipramina (blochează absorbția norepinefrinei de către fibrele nervoase), amfetamina (promovează simultan eliberarea norepinefrinei și blochează absorbția acesteia), inhibitori MAO (suprimă metabolismul catecolaminelor), etc. În acest sens, a apărut ipoteza catecolaminei a stărilor depresive, conform căreia depresie psihică asociat cu o lipsă de catecolamine în creier.
La începutul anilor '50, farmacologii au descoperit că binecunoscutul halucinogen, acidul lisergic dietilamina (LSD), nu numai că era similar ca structură chimică cu serotonina, dar a neutralizat și o parte din acesta. efecte farmacologice(prin blocarea receptorilor serotoninei). Prin urmare, s-a sugerat că tulburările în metabolismul serotoninei pot fi o cauză a unor boli mintale specifice.
Ei cred că sunt antipsihotice, precum aminazina (clorpromazina) și haloperidolul, prin îmbunătățirea sintezei catecolaminelor, sunt capabile să blocheze receptorii de dopamină din creier.
PEPTIDE ȘI REACȚII DURERE
În anii 1970, în creierul diferitelor vertebrate au fost descoperiți receptori specifici de morfină. Acești receptori sunt concentrați pe membranele sinaptice, cele mai bogate în ele Sistemul limbic, de care depinde răspunsul emoțional. Ulterior, peptidele endogene au fost izolate din țesutul cerebral care imită diferitele efecte ale morfinei atunci când sunt injectate. Aceste peptide, care au capacitatea de a se lega în mod specific de receptorii de opiacee, se numesc endorfine și encefaline.
S-a dovedit că peptidele cu activitate asemănătoare morfinei sunt derivați ai hormonului β-lipotrop al glandei pituitare. S-a stabilit că β-endorfină este un fragment de β-lipotropină de la 61 la 91, y-endorfină de la 61 la 77 și α-endorfină de la 61 la 76 de resturi de aminoacizi.
Enkefalinele sunt, de asemenea, fragmente de β-lipotropină, dar sunt mult mai mici decât endorfinele. Enkefalinele sunt pentapeptide. Cele două pentapeptide cele mai studiate sunt metionin encefalina (Tyr-Gly-Gly-Phen-Met) și leucină-encefalina (Tir-Gly-Gly-Phen-Leu). Conținutul de encefaline de metionină din creier este de 4 ori mai mare decât conținutul de encefaline de leucină.
LICHID CORESPINAL
Volum total fluid cerebrospinal(lichidul cefalorahidian) în mod normal la un adult este de aproximativ 125 ml, care se reînnoiește la fiecare 3-4 ore. Lichiorul este uneori considerat ca un transudat primar sau ultrafiltrat de plasmă. Compoziția lichidului cefalorahidian diferă semnificativ de compoziția plasmei sanguine, ceea ce ne permite să atribuim rolul principal în implementarea funcției de barieră endoteliului vascular din sistemul nervos. Apa din lichior reprezintă 99%, reziduul dens reprezintă aproximativ 1%. Compoziția chimică a lichidului cefalorahidian este prezentată în tabel. 58.
Conținutul de proteine din lichidul cefalorahidian este nesemnificativ (0,15-0,40 g/l), iar raportul albumină/globulină este de 4; lipidele sunt de sute de ori mai puține decât în plasma sanguină. Este posibil să nu existe deloc lipide plasmatice în lichidul cefalorahidian. Conținutul total de substanțe care conțin azot cu greutate moleculară mică, în special aminoacizi, este de 2-2,5 ori mai mic decât în sânge. În țesutul cerebral, așa cum sa menționat deja, cantitatea de aminoacizi liberi este mare și de multe ori depășește concentrația acestora în sânge și cu atât mai mult în lichidul cefalorahidian. S-a stabilit că unii aminoacizi (de exemplu, acidul glutamic) pătrund cu greu în bariera hemato-encefalică. În același timp, amidele de aminoacizi (în special, glutamina) depășesc cu ușurință această barieră. Conținutul de glucoză în lichidul cefalorahidian este relativ mare (2,50-4,16 mmol/l), dar ceva mai mic decât în sânge, iar concentrația de glucoză în lichidul cefalorahidian poate crește sau scădea în funcție de modificările nivelului de glucoză din sânge.
În ceea ce privește conținutul de sodiu și potasiu, lichidul cefalorahidian nu este practic diferit de plasma sanguină. Calciul din lichidul cefalorahidian este de aproape 2 ori mai mic decât în plasma sanguină. Conținutul de clor este vizibil mai mare, iar concentrația ionilor de bicarbonat este puțin mai mică în lichidul cefalorahidian decât în plasmă. Prin urmare, compozitia minerala lichidul cefalorahidian are, de asemenea, trăsături caracteristice în comparație cu cel din plasma sanguină. Toate acestea dau motive să credem că pătrunderea substanțelor prin membrana endoteliului vascular al sistemului nervos este un activ. proces biochimic. Sursele de energie pentru transportul activ sunt procesul oxidare aerobă glucoza si numai intr-o mica masura glicoliza.
Examinarea lichidului cefalorahidian stări patologice are important semnificație clinică. S-a stabilit că în acută meningita purulenta continutul de proteine din lichidul cefalorahidian poate creste brusc (5-20 g/l) fata de norma (0,15-0,40 g/l). De asemenea, concentrația de glucoză în lichidul cefalorahidian se modifică semnificativ. Hipoglicorahia (scăderea conținutului de zahăr în lichidul cefalorahidian) este caracteristică meningitei, în timp ce hiperglicorahia (conținutul crescut de zahăr în lichidul cefalorahidian) se observă în encefalită, diabet, etc. Caracteristica este o scădere a concentrației de clor în lichidul cefalorahidian cu meningită. și o creștere a conținutului său cu encefalită. De asemenea, s-a demonstrat că în caz de meningită, accident vascular cerebral, tumori cerebrale, leziuni, activitatea aspartat aminotransferazei, lactat dehidrogenazei și o serie de alte enzime crește în lichidul cefalorahidian.



