Neuronul este o celulă nervoasă. Bazele teoriei neuronale
În general, în funcție de sarcinile și responsabilitățile atribuite neuronilor, aceștia sunt împărțiți în trei categorii:
- Neuroni senzoriali primiți și transmiteți impulsuri de la receptori „în centru”, adică sistem nervos central. În plus, receptorii înșiși sunt celule special antrenate ale organelor senzoriale, mușchilor, pielii și articulațiilor care pot detecta modificări fizice sau chimice în interiorul și în afara corpului nostru, le pot transforma în impulsuri și le pot transmite cu bucurie neuronilor senzoriali. Astfel, semnalele circulă de la periferie la centru.
Următorul tip:
- Neuroni motori (motorii), care toarcă, fircha și bip, poartă semnale venite din creier sau măduva spinării, la organele executive, care sunt mușchii, glandele etc. Da, asta înseamnă că semnalele merg de la centru la periferie.
bine si neuroni intermediari (intercalari), pur și simplu, sunt „prelungitoare”, adică. primesc semnale de la neuronii senzoriali și trimit aceste impulsuri mai departe către alți neuroni intermediari sau direct către neuronii motori.
În general, așa se întâmplă: în neuronii senzoriali, dendritele sunt conectate la receptori, iar axonii sunt conectați la alți neuroni (interneuroni). În neuronii motori, dimpotrivă, dendritele sunt conectate la alți neuroni (interneuroni), iar axonii sunt conectați la un anumit efector, adică. un stimulator al contracției musculare sau al secreției glandelor. Ei bine, în consecință, interneuroni iar dendritele și axonii se conectează la alți neuroni.
Se pare că cea mai simplă cale pe care poate parcurge un impuls nervos va consta din trei neuroni: unul senzorial, unul intercalar și unul motor.
Da, acum să ne amintim de tip - un foarte „patolog nervos”, cu un zâmbet rău intenționat, bătând în genunchi cu ciocanul său „magic”. Suna familiar? Iată-l cel mai simplu reflex: când loveşte tendonul rotulian, mușchiul atașat acestuia se întinde și semnalul de la celulele senzitive (receptorii) situate în el este transmis prin neuronii senzoriali la măduva spinării. Și deja în ea, neuronii senzoriali contactează fie prin intercalare, fie direct cu neuronii motori, care ca răspuns trimit impulsuri înapoi aceluiași mușchi, făcându-l să se contracte și piciorul să se îndrepte.
Măduva spinării în sine este amplasată convenabil în coloana vertebrală. Este moale și vulnerabil, motiv pentru care se ascunde în vertebre. Măduva spinării are o lungime de doar 40-45 de centimetri, groasă cât un deget mic (aproximativ 8 mm) și cântărește aproximativ 30 de grame! Dar, în ciuda tuturor fragilității sale, măduva spinării este centrul de control al unei rețele complexe de nervi răspândite în tot corpul. Aproape ca un centru de control al misiunii! :) Fara el SIstemul musculoscheletal, nici de bază organe vitale Ei bine, pur și simplu nu pot acționa și lucra.
Măduva spinării își are originea la nivelul marginii foramenului occipital al craniului și se termină la nivelul primei și celei de-a doua vertebre lombare. Dar sub măduva spinării în canalul rahidian există un mănunchi atât de dens de rădăcini nervoase, amuzant numit cauda equina, aparent pentru asemănarea cu acesta. Asa de, coadă de cal- Aceasta este o continuare a nervilor care ies din măduva spinării. Ei sunt responsabili de inervație membrele inferioareși organele pelvine, adică transmite semnale de la măduva spinării către ei.
Măduva spinării este înconjurată de trei membrane: moale, arahnoidă și tare. Și spațiul dintre membranele moi și arahnoidian este de asemenea umplut o cantitate mare fluid cerebrospinal. Prin foramenele intervertebrale, nervii spinali pleacă din măduva spinării: 8 perechi de cervicali, 12 toracici, 5 lombari, 5 sacrali și 1 sau 2 coccigieni. De ce abur? Da, pentru că nervul spinal iese prin două rădăcini: posterior (sensibil) și anterior (motor), conectate într-un singur trunchi. Deci, fiecare astfel de pereche controlează o anumită parte a corpului. Adică, de exemplu, dacă ați apucat din greșeală o tigaie fierbinte (Doamne ferește! Pah-pah-pah!), atunci un semnal de durere apare imediat la terminațiile nervului senzorial, intrând imediat în măduva spinării și de acolo - în se dublează nervul motor, care transmite ordinul: „Akhtung-akhtung! Scoate-ți mâna imediat!” Mai mult, credeți-mă, acest lucru se întâmplă foarte repede - chiar înainte ca creierul să înregistreze impulsul durerii. Drept urmare, reușiți să trageți mâna departe de tigaie înainte de a simți durere. Desigur, această reacție ne salvează de arsuri grave sau alte daune.
În general, aproape toate noastre automate și actiuni reflexe sunt controlate de măduva spinării, ei bine, cu excepția celor care sunt monitorizate de creierul însuși. Ei bine, de exemplu: percepem ceea ce vedem cu ajutorul lui nervul optic mergând în creier și, în același timp, ne întoarcem privirea în direcții diferite folosind muschii ochilor, care sunt deja controlate de măduva spinării. Da, și plângem la fel la ordinele măduvei spinării, care „gestionează” glandele lacrimale.
Putem spune că acțiunile noastre conștiente provin din creier, dar de îndată ce începem să efectuăm aceste acțiuni automat și reflex, ele sunt transferate în măduva spinării. Deci, atunci când doar învățăm să facem ceva, atunci, desigur, ne gândim în mod conștient, gândim și înțelegem fiecare mișcare, ceea ce înseamnă că folosim creierul, dar în timp o putem face deja automat și asta înseamnă că creierul transferă „frâiele puterii” acestei acțiuni pe cea spinală, doar că deja s-a plictisit și neinteresant... pentru că creierul nostru este foarte curios, curios și adoră să învețe!
Ei bine, este timpul să fim curioși......
Funcţie sistem nervos este
1) managementul activităților diferitelor sisteme care alcătuiesc întregul organism,
2) coordonarea proceselor care au loc în ea,
3) stabilirea de relaţii între organism şi mediul extern.
Activitatea sistemului nervos este de natură reflexivă. Reflexul (lat. reflexus - reflectat) este răspunsul organismului la orice impact. Aceasta poate fi influență externă sau internă (din mediul extern sau din propriul corp).
Unitatea structurală și funcțională a sistemului nervos este neuron(celula nervoasa, neurocit). Un neuron este format din două părți - corpȘi proceselor. Procesele unui neuron, la rândul lor, sunt de două tipuri - dendriteȘi axonii. Procesele de-a lungul cărora impulsul nervos este transportat către corpul celulei nervoase sunt numite dendrite. Procesul prin care impulsul nervos este direcționat de la corpul neuronului către o altă celulă nervoasă sau către țesutul de lucru se numește axon. Nervcelulăcapabil să transmită nervosimpuls într-o singură direcțienii - de la dendrite prin corpul celular laaxon.
Neuronii din sistemul nervos formează lanțuri de-a lungul cărora sunt transmise (deplasate) impulsurile nervoase. Transmiterea unui impuls nervos de la un neuron la altul are loc la locurile de contact și este asigurată de un tip special de structuri anatomice numite sinapsa interneuronalebufnițe.
Într-un lanț nervos, diferiți neuroni îndeplinesc diferite funcții. În acest sens, se disting următoarele trei tipuri principale de neuroni:
1. neuron senzorial (aferent)..
2. interneuron.
3. neuron efector (eferent)..
sensibil, (receptiv,sauaferente) neuronii. Principalele caracteristici ale neuronilor senzoriali:
A) Ta mâncat neuroni senzoriali se află întotdeauna în noduri (ganglioni spinali), în afara creierului sau măduvei spinării;
b) un neuron senzorial are două procese - unul dendrit și unul axon;
V) dendrita neuronului senzorial urmează la periferie până la unul sau altul organ și se termină acolo cu un final sensibil - receptor. Receptor acesta este un organ care este capabil să transforme energia influenței externe (iritarea) într-un impuls nervos;
G) axonul neuronului senzorial trimis la sistemul nervos central, la măduva spinării sau la trunchiul cerebral, ca parte a rădăcinilor dorsale a nervilor spinali sau a nervilor cranieni corespunzători.
Un receptor este un organ care este capabil să transforme energia influenței externe (iritarea) într-un impuls nervos. Este situat la capătul dendritei neuronului senzorial
Se disting următoarele: tipuri de retetetori in functie de locatie:
1) Exteroceptori percepe iritația din mediul extern. Sunt localizate în tegumentul exterior al corpului, în piele și mucoase, în organele senzoriale;
2) Interoceptori fi iritat de mediu intern organism, sunt localizate în organele interne;
3) Proprioceptori percepe iritații din sistemul musculo-scheletic (în mușchi, tendoane, ligamente, fascie, capsule articulare.
Funcția neuronului senzorial– perceperea unui impuls de la receptor și transmiterea acestuia către sistemul nervos central. I.P Pavlov a atribuit acest fenomen începutului procesului de analiză.
Insertional, (neuron asociativ, de închidere sau conductor ) realizează transferul excitaţiei de la neuronul senzitiv (aferent) la cei eferenti. Neuronii de închidere (intercalari) se află în sistemul nervos central.
Efector, (eferent)neuron. Există două tipuri de neuroni eferenți. Acest dvineuron gator,Șineuron secretor. Proprietăți de bază neuroni motorii:
Corpurile celulare ale neuronilor motori sunt localizate în sistemul nervos central, în coarnele anterioare ale măduvei spinării.
Axonii neuronilor motori sunt trimiși ca parte a fibrelor nervoase către organele de lucru (mușchii striați ai sistemului musculo-scheletic).
Proprietăți de bază neuroni secretori:
corpurile neuronilor secretori sunt localizate în nodulii simpatici și parasimpatici;
axonii neuronilor secretori sunt direcționați către organele interne.
Principiul principal al sistemului nervos este principiul unui răspuns reflex la iritare.
În conformitate cu aceasta, structura sistemului nervos se bazează pe arcul reflex. Arc reflex este un lanț de celule nervoase de-a lungul căruia un impuls nervos se deplasează de la locul său de origine (de la receptor) la organul de lucru (la efector).
Cel mai simplu arc reflex(Fig. 184) este format din doar doi neuroni - senzitiv și motor (aferent și efector). Corpul primului neuron (sensibil) este situat în ganglionul spinal. Procesul periferic al acestei celule se încheie cu un receptor care percepe iritația. Receptorul transformă această iritație într-un impuls nervos. Impulsul nervos ajunge în corpul celulei nervoase de-a lungul dendritei și apoi se deplasează de-a lungul axonului până la măduva spinării.
ÎN substanța cenușie a măduvei spinăriicreier acest proces al celulelor sensibile se formează sinapsa cu corpul celui de-al doilea neuron (motor). La sinapsa interneuronă, excitația nervoasă este transferată de la un neuron sensibil (aferent) la un neuron motor (eferent). Proces neuron motor iese din măduva spinării ca parte a rădăcinilor ventrale nervi spinaliși este direcționat către organul de lucru, controlând munca mușchiului.
De regulă, arcul reflex nu este format din doi neuroni, ci este mult mai complex. Între doi neuroni - receptor(aferent) și efector(eferent) - există unul sau mai multe intercalar(închiderea) neuronilor. În acest caz, excitația de la neuronul receptor de-a lungul procesului său central este transmisă nu direct la celula nervoasă efectoră, ci la unul sau mai mulți interneuroni. Rolul interneuronilor în măduva spinării este îndeplinit de celulele aflate în interior materie cenusie stâlpii din spate. Iritația chiar și a celui mai mic număr de receptori poate fi transmisă nu numai unui anumit segment al măduvei spinării, ci și răspândită la celulele mai multor segmente învecinate. Ca urmare a acestui fapt, răspunsul este o contracție nu a unui mușchi sau chiar a unui grup de mușchi, ci a mai multor grupuri deodată. Deci, ca răspuns la iritare, apare o mișcare complexă, reflexă. Acesta este unul dintre răspunsurile corpului (reflex) ca răspuns la iritare.
Marele merit al lui I.P Pavlov este că a extins doctrina reflexului la întregul sistem nervos, pornind de la secțiunile cele mai de jos și terminând cu secțiunile sale cele mai înalte, și a demonstrat experimental natura reflexă a tuturor formelor de activitate vitală a corpului. Potrivit lui I.P. forma simpla a deyactivitatea sistemului nervos, trebuie safi desemnat ca un ref neconditionatlex. Reflexul necondiționat este o formă constantă de activitate a sistemului nervos, înnăscută, cu trăsături caracteristice fiecărui tip.
În plus, există legături temporare cu mediul dobândite în timpul vieții unui individ. Capacitatea de a dobândi conexiuni temporare permite organismului să stabilească cele mai diverse și complexe relații cu mediul extern. I. P. Pavlov a numit această formă de activitate reflexă reflex condiționat(spre deosebire de non-reflex). Locul de închidere a reflexelor condiționate este scoarța cerebrală. Creierul și cortexul său sunt baza activității nervoase superioare.
Sistemul nervos uman este împărțit în mod convențional, conform principiului topografic, în două părți - central și periferic.
Sistemul nervos central include măduva spinăriiȘi creier. Măduva spinării și creierul sunt formate din substanță cenușie și albă.
Substanța cenușie a coloanei vertebraleși creierul este o colecție de corpuri de celule nervoase. materie albă- acestea sunt fibre nervoase, procese ale celulelor nervoase. Fibrele nervoase formează căile măduvei spinării și creierului și conectează diferite părți ale sistemului nervos central și diferiți nuclei ( centrii nervosi) între ei.
Sistem nervos periferic constă din rădăcini, nervi, ramurile lor, plexuri și noduri aflate în ele diverse departamente corpul uman.
Conform unei alte clasificări anatomice și funcționale, sistemul nervos unificat este, de asemenea, împărțit convențional în două părți: I) somatic și 2) vegetativ.
Samatisistemul nervos logic asigură inervație în principal organismului - soma, și anume pielii și sistemului musculo-scheletic.
Nervos autonom (autonom).sistem inervează totul organe interneși reglează procesele metabolice în toate organele și țesuturile.
Sistemul nervos autonom este, la rândul său, împărțit în două părți: parasimpatic Și simpatic. În fiecare dintre aceste părți, ca și în sistemul nervos somatic, există secțiuni centrale și periferice.
Această diviziune a sistemului nervos, în ciuda convenționalității sale, s-a dezvoltat în mod tradițional și pare destul de convenabilă pentru studierea sistemului nervos ca întreg și a părților sale individuale. În acest sens, în viitor vom adera la această clasificare și în prezentarea materialului.
Neuron(neuron grecesc - nerv) - celula nervoasa, format dintr-un corp și procese care se extind din acesta - dendrite relativ scurte și un axon lung; unitatea structurală și funcțională de bază a sistemului nervos. Neuronii primesc impulsuri nervoase de la receptorii din sistemul nervos central (nervul senzitiv) și generează impulsuri transmise de la sistemul nervos central către organele executive (nervul motor). Aceste N. sunt conectate între ele prin alte celule nervoase (N. intercalar). N. interacționează între ele și cu celulele organe executive prin sinapse. La rotifere, numărul H este 102, la om este mai mare de 1010.
Elementele structurale și funcționale ale unei celule nervoase. În fiecare celulă nervoasă se pot distinge patru elemente principale: corpul, sau soma, dendrite, axonul și terminalul presinaptic al axonului. Fiecare dintre aceste elemente îndeplinește o funcție specifică. Corpul neuronal contine diverse organite intracelulare necesare asigurarii activitatii vitale a intregii celule: nucleu, ribozomi, reticul endoplasmatic, complex lamelar (aparatul Golgi), mitocondrii. Aici are loc sinteza principală a macromoleculelor, care pot fi apoi transportate în dendrite și axon. Membrana corpului majorității neuronilor este acoperită cu sinapse și astfel se joacă rol importantîn perceperea şi integrarea semnalelor provenite de la alţi neuroni.
Dendritele și axonii provin din corpul celular. În cele mai multe cazuri, dendritele sunt foarte ramificate. Ca urmare, suprafața lor totală depășește semnificativ suprafața corpului celular. Acest lucru creează condiții pentru plasarea pe dendrite cantitati mari sinapsele. Astfel, dendritele joacă un rol principal în percepția informațiilor neuronale. Membrana dendritelor, ca și membrana corpului neuronilor, conține un număr semnificativ de molecule de proteine care funcționează ca receptori chimici cu sensibilitate specifică la anumite substanțe chimice. Aceste substanțe sunt implicate în transmiterea semnalelor de la celulă la celulă și sunt mediatori ai excitației și inhibiției sinaptice. Funcția principală a axonului este de a conduce un impuls nervos - un potențial de acțiune. Capacitatea unui potențial de acțiune de a se propaga fără slăbire oferă implementare eficientă semnal pe toată lungimea axonului, care în unele celule nervoase atinge multe zeci de centimetri. Astfel, sarcina principală a axonului este de a transporta semnale pe distanțe lungi, conectând celulele nervoase între ele și cu organele executive.
Terminalul axonal este specializat în transmiterea unui semnal către alți neuroni (sau celule ale organelor executive). Prin urmare, conține organele speciale: vezicule sinaptice sau vezicule care conțin mediatori chimici. Membrana terminațiilor presinaptice ale axonului, spre deosebire de axonul însuși, este echipată cu receptori specifici care pot răspunde la diferiți mediatori.
Definiții, semnificații ale cuvintelor din alte dicționare:
Dicţionar filosofic
(din grecescul neuron - nerv) - o celulă nervoasă formată dintr-un corp și procese care se extind din acesta - dendrite relativ scurte și un axon lung; unitatea structurală și funcțională de bază a sistemului nervos. Conducem impulsurile nervoase de la receptori la sistemul nervos central...
(celula nervoasa) - principala unitate structurala si functionala a sistemului nervos; un neuron generează, primește și transmite impulsuri nervoase, transmițând astfel informații dintr-o parte a corpului în alta (vezi figura). Fiecare neuron are un corp celular mare (sau pericarion (...
Enciclopedie psihologică
Celula nervoasă, principala unitate structurală și funcțională a sistemului nervos. Deși au o mare varietate de forme și dimensiuni și sunt implicați într-o gamă largă de funcții, toți neuronii constau dintr-un corp celular, sau soma, care conține un nucleu și procese nervoase: axonul și...
Țesutul nervos este format din două tipuri de celule diferite din punct de vedere morfologic și funcțional. Una dintre ele include celulele nervoase sau neuronii propriu-zis, iar celălalt include celulele neurogliale sau pur și simplu glia. Ambele provin dintr-o populație comună de celule progenitoare care există numai pe stadiu timpuriu Dezvoltarea embrionară creier. În procesul de diferențiere, acești doi tipul de celule sunt izolați și apoi se specializează în îndeplinirea diferitelor sarcini.
Calea spre studierea morfologiei i.e. În aparență, neuronii au fost descoperiți pentru prima dată la sfârșitul anilor 60 - începutul anilor 70 ai secolului al XIX-lea de către medicul și anatomistul italian Camillo Golgi (Golgi S.). Într-o zi, a decis să folosească nitrat de argint pentru a colora preparatele din creier și a descoperit că această substanță este absorbită selectiv de celulele nervoase. Această metodă a fost ulterior îmbunătățită de celebrul histolog spaniol Santiago Ramon y Cajal S. și, folosindu-l, a obținut date care au făcut posibilă crearea bazei. teoria neuronală. În 1906, Golgi și Ramon-Cajal au fost premiați Premiul Nobelîn fiziologie și medicină „pentru lucrări privind studiul structurii sistemului nervos”.
La sfârșitul secolului al XVIII-lea, italianul Luigi Galvani (Galvani L.) a descoperit că mușchii și celulele nervoase ale animalelor produc electricitate. La mijlocul secolului al XIX-lea în Germania, Emile DuBois-Reymond E. dispunea deja de tehnologie suficient de avansată pentru a înregistra semnale bioelectrice într-o fibră nervoasă. Hermann Helmholtz (von Helmholtz N.) în 1854 a fost capabil să măsoare viteza de trecere impulsuri nervoase.
De atunci s-a făcut o cantitate mare studii ale structurii și funcției neuronilor, care au făcut posibilă formularea fundamentelor teoriei neuronale.
Dintre 1011 neuroni creier uman au fost descoperite multe celule care sunt diferite între ele, dar în structura lor se pot găsi de obicei comune Caracteristici(Fig. 3.1). Fiecare neuron are un corp (alte denumiri pentru această parte a neuronului: soma, perikarion), care conține un nucleu și organele citoplasmatice, unde are loc sinteza proteinelor, neurotransmițătorilor și altele. componente importante viata celulara. Când corpul este distrus, întreaga celulă moare inevitabil.
Două tipuri de procese se extind din corpul celular, care sunt fire subțiri de citoplasmă; se numesc dendrite si axon. Numărul de dendrite din diferite celule poate varia semnificativ, majoritatea neuronilor au destul de mulți dintre ele, iar fiecare dintre dendrite se ramifică ca un copac, iar numeroasele sale ramuri sunt concepute pentru a primi semnale transmise de celulele învecinate. După ce au primit astfel de semnale, dendritele le conduc către corpul celular.
Spre deosebire de numărul variabil de dendrite, orice celulă nervoasă poate avea un singur axon, care conduce semnalele electrice într-o singură direcție: departe de corpul celular. Aceste semnale electrice sunt numite potențiale de acțiune și au o amplitudine de aproximativ 100 milivolți (mV - miimi de volt) și o durată de aproximativ 1 milisecundă (ms - miimi de secundă). Potențialele de acțiune apar de obicei la dealul axonului, locul în care axonul provine din soma și se propagă de-a lungul axonului la viteze cuprinse între 1 și 100 m/s fără a modifica amplitudinea semnalului.
Diametrul axonului este aproape același pe toată lungimea sa, celule diferite valoarea sa variază de la 0,2 la 20 µm. Această împrejurare afectează viteza de transmitere a semnalului: cu cât axonul este mai gros, cu atât potenţialul de acţiune este condus mai repede de-a lungul acestuia. Lungimea axonilor din diferite celule poate varia foarte mult: de la 0,1 mm la 1 m (și la unele specii de animale - până la 3 m). Mulți axoni sunt închiși într-o teacă specială formată prin procesele anumitor celule gliale. Acest caz este format din mielină, o substanță asemănătoare grăsimii cu proprietățile unui izolator electric: în zona învelișului de mielină, nu apar semnale electrice.
Teaca de mielină a axonului este întreruptă în mod regulat de zone fără mielină - acestea se numesc noduri de Ranvier. Potențialele de acțiune se propagă de-a lungul acestor interceptări, de parcă sărind prin zonele mielinizate de la o interceptare la alta (acest tip de transmisie a semnalului se numește saltator, din latinescul saltare - a sări), deci viteza de conducere este destul de mare. Unii axoni nu au un înveliș de mielină: spre deosebire de fibrele mielinice, ele sunt numiți nemielinizați (conform altei terminologii, fibrele mielinice și nemielinice se disting ca pulpoase și nemielinizate). Potențialele de acțiune călătoresc mai lent de-a lungul fibrelor nemielinice: aici nu „sar”, ci „se târăște” pe toată lungimea axonului.
Natura potențialelor de acțiune, cu ajutorul cărora informația este transmisă în sisteme senzoriale, motorii sau motivaționale, este aceeași, iar potențialele de acțiune în sine sunt stereotipe. Apariția și implementarea lor pot fi înregistrate folosind dispozitive speciale, dar din natura unei astfel de înregistrări este imposibil să se judece ce fel de informații transmit potențialele de acțiune înregistrate: mirosul de liliac înflorit sau verdele smarald mângâietor al gazonului tuns sau extensia piciorul dreptîn genunchi sau saliva. Conținutul informațiilor transmise este determinat nu de forma potențialelor de acțiune, ci de asocierile specifice ale neuronilor într-unul sau altul sistem senzorial, motor sau motivațional: fiecare tip de informație este transmis folosind potențiale de acțiune stereotipe, dar de-a lungul propriei căi neuronale.
Aproape de terminațiile lor, majoritatea axonilor se împart în ramuri colaterale subțiri sau terminale axonale, iar unii dintre ei se pot întoarce chiar înapoi - acestea sunt colaterale inverse. Terminalele axonale intră în contact cu alte celule, cel mai adesea cu dendritele lor, mai rar cu corpul și chiar mai rar cu axonul. Axonii neuronilor eferenți contactează celulele organelor de lucru, care sunt mușchi sau glande exocrine. Zona de contact dintre două celule se numește sinapsă. În conformitate cu acest termen, celula care transmite semnalul este numită presinaptică, iar celula care primește semnalul este numită postsinaptică. În marea majoritate a cazurilor, aceste celule nu sunt conectate anatomic și între ele există o despicatură sinaptică, care este umplută cu un lichid asemănător cu plasma sanguină în compoziția sa (sinapsele electrice reprezintă un tip special de contacte intercelulare - vezi capitolul 5).
Datorită separării anatomice, o celulă presinaptică poate influența o celulă postsinaptică doar cu ajutorul unui intermediar chimic - un neurotransmițător sau neurotransmițător. Transmițătorul trebuie eliberat din terminalul axon al celulei presinaptice atunci când un potențial de acțiune se apropie de acest terminal.
Pe baza numărului de procese citoplasmatice, se obișnuiește să se facă distincția între neuronii unipolari, bipolari și multipolari. Neuronii unipolari au un singur neuroni, de obicei foarte ramificat proces primar. Una dintre ramurile sale funcționează ca axon, iar restul ca dendrite. Astfel de celule se găsesc adesea în sistemul nervos al nevertebratelor, dar la vertebrate se găsesc numai în unele ganglioni ai sistemului nervos autonom.
Celulele bipolare au două procese (Fig. 3.2): dendrita transportă semnale de la periferie către corpul celular, iar axonul transmite informația din corpul celular către alți neuroni. Așa arată, de exemplu, unii neuroni senzoriali, aflați în retina ochiului și în epiteliul olfactiv.

Același tip de neuroni ar trebui să includă și celulele sensibile ale ganglionilor spinali, care percep, de exemplu, atingerea pielii sau durerea, deși formal un singur proces se extinde din corpul lor, care este împărțit în ramuri centrale și periferice. Astfel de celule sunt numite pseudounipolare, s-au format inițial ca neuroni bipolari, dar în timpul dezvoltării, cele două procese ale lor s-au unit într-unul, în care o ramură funcționează ca un axon și cealaltă ca o dendră.
Celulele multipolare au un singur axon, dar pot exista o mulțime de dendrite care se extind din corpul celular și apoi se divid în mod repetat, formând numeroase sinapse pe ramurile lor cu alți neuroni. De exemplu, aproximativ 8.000 de sinapse sunt formate pe dendritele unui singur motor neuron al măduvei spinării și până la 150.000 de sinapse pot fi formate pe dendritele celulelor Purkinje situate în cortexul cerebelos. Neuronii Purkinje sunt, de asemenea, cei mai mulți celule mari creierul uman: diametrul corpului lor este de aproximativ 80 de microni. Și lângă ele sunt celule granulare minuscule, diametrul lor este de doar 6-8 microni. Neuronii multipolari se găsesc cel mai adesea în sistemul nervos și printre ei sunt identificate multe celule care nu sunt asemănătoare ca aspect între ele.
Neuronii sunt de obicei clasificați nu numai după forma lor, ci și după funcția lor și după locul lor în lanțul de celule care interacționează. Unele dintre ele au terminații sensibile speciale - receptori, care sunt excitați atunci când sunt expuși la orice factori fizici sau chimici, cum ar fi, de exemplu, lumina, presiunea sau adăugarea anumitor molecule. După excitarea receptorilor, neuronii senzoriali transmit informații către sistemul nervos central, adică. conduc semnalele centripet sau aferent (latin afferens - aducere).
Un alt tip de celulă transmite comenzi de la sistemul nervos central către mușchii scheletici sau netezi, către mușchiul inimii sau către glandele exocrine. Aceștia sunt fie neuroni motori, fie autonomi, prin care semnalele se propagă centrifug, iar astfel de neuroni înșiși sunt numiți eferenti (lat. efferens - eferent).
Toți ceilalți neuroni aparțin categoriei de interneuroni sau interneuroni, care formează cea mai mare parte a sistemului nervos - 99,98% din numărul total celule. Printre aceștia se numără, așa cum sa menționat deja în capitolul 2, neuronii locali și de proiecție. Un alt nume pentru neuronii de proiecție este neuronii releu; au de obicei axoni lungi, cu ajutorul cărora aceste celule pot transmite informații procesate în regiuni îndepărtate ale creierului. Interneuronii locali au axoni scurti, aceste celule procesează informații în circuite locale limitate și interacționează în primul rând cu neuronii vecini.
Ramon-Cajal a formulat, de asemenea, două principii care au stat la baza teoriei neuronale și și-au păstrat semnificația până astăzi:
1. Principiul polarizării dinamice. Înseamnă că semnalul electric călătorește prin neuron într-o singură direcție și previzibilă.
2. Principiul specificității compusului. În conformitate cu acest principiu, neuronii nu vin în contact aleatoriu, ci doar cu anumite celule țintă, iar citoplasma celulelor în contact nu se conectează și se menține întotdeauna un decalaj sinaptic între ele.
Versiunea modernă a teoriei neuronale conectează anumite părți ale celulei nervoase cu natura semnalelor electrice care apar în ele. Un neuron tipic are patru regiuni definite morfologic: dendrite, soma, axon și terminal axon presinaptic. Când un neuron este excitat, în el apar secvenţial patru tipuri de semnale electrice: de intrare, combinate, conductoare şi de ieşire (Fig. 3.3). Fiecare dintre aceste semnale apare doar într-o anumită regiune morfologică.

Pentru a înțelege diferențele dintre aceste semnale, trebuie să înțelegeți natura impulsurilor nervoase. Din afară și interior Membrana plasmatică a unui neuron conține diferite sarcini electrice: pozitive la exterior și negative la interior. Diferența dintre ele se numește potențial de membrană de repaus. Dacă considerăm sarcina externă egală cu zero, atunci diferența de încărcături dintre cele externe și suprafete interioareîn majoritatea neuronilor se dovedește a fi aproape de -65 mV, deși poate celule individuale variază de la -40 la -80 mV.
Apariția acestei diferențe de sarcină se datorează distribuției inegale a ionilor de potasiu, sodiu și clor în interiorul și în afara celulei, precum și permeabilitatea mai mare a celulei de repaus. membrana celulara numai pentru ionii de potasiu (vezi capitolul 4).
În celulele excitabile, care includ celulele nervoase și musculare, potențialul de repaus poate varia foarte mult, iar această capacitate stă la baza apariției semnalelor electrice. O scădere a potențialului de repaus, de exemplu, de la -65 la -60 mV, se numește depolarizare, iar o creștere, de exemplu, de la -65 la -70 mV, se numește hiperpolarizare.
Dacă depolarizarea atinge un anumit nivel critic, de exemplu -55 mV, atunci permeabilitatea membranei pentru ionii de sodiu va scădea un timp scurt devine maximă, se repezi în celulă și, în legătură cu aceasta, diferența de potențial transmembranar scade rapid la 0 și apoi capătă valoare pozitivă. Această împrejurare duce la închidere canale de sodiuși eliberarea rapidă a ionilor de potasiu din celulă prin canale destinate numai acestora: ca urmare, valoarea inițială este restabilită potențial de membrană. Aceste modificări care apar rapid în potențialul membranei sunt numite potențiale de acțiune. Potențialul de acțiune este un semnal electric conductiv, se răspândește rapid de-a lungul membranei axonului până la capătul său și nu își schimbă amplitudinea nicăieri.
Pe lângă potențialele de acțiune dintr-o celulă nervoasă, din cauza modificărilor permeabilității membranei acesteia, pot apărea semnale locale sau locale: potențial receptor și potențial postsinaptic. Amplitudinea lor este semnificativ mai mică decât cea a potențialului de acțiune în plus, scade semnificativ pe măsură ce semnalul se propagă. Din acest motiv, potențialele locale nu se pot propaga prin membrană departe de punctul lor de origine.
3.4. Semnale de intrare
Semnalele de intrare sunt fie receptor, fie potenţial postsinaptic. Un potențial receptor se formează în terminațiile unui neuron senzitiv atunci când un anumit stimul acționează asupra lor: întindere, presiune, lumină, Substanta chimicași așa mai departe. Acţiunea stimulului determină deschiderea unor canale ionice în membrană, iar fluxul ulterior de ioni prin aceste canale modifică valoarea iniţială a potenţialului de repaus; în majoritatea cazurilor apare depolarizarea. Această depolarizare este potențialul receptor, amplitudinea sa este proporțională cu puterea stimulului curent.
Potențialul receptor se poate răspândi de la locul stimulului de-a lungul membranei, dar, de regulă, pe o distanță relativ scurtă. Cert este că amplitudinea potențialului receptor scade odată cu distanța de la locul de acțiune al stimulului și la o distanță de numai 1 mm de acest loc amplitudinea potențialului receptor este de numai 1/3 din valoarea inițială, iar după încă 1 mm deplasarea depolarizantă va dispărea complet.
Al doilea tip de semnal de intrare este potențialul postsinaptic. Se formează pe celula postsinaptică după ce celula presinaptică excitată trimite un curier chimic special pentru ea - un neurotransmițător. Ajuns la celula postsinaptică prin difuzie, mediatorul se atașează de proteinele receptorului specific din membrana sa (vezi capitolul 1), ceea ce determină deschiderea canalelor ionice. Curentul ionic rezultat prin membrana postsinaptică modifică valoarea inițială a potențialului de repaus - această schimbare este potențialul postsinaptic.
În unele sinapse, o astfel de schimbare reprezintă depolarizare și, dacă atinge un nivel critic, neuronul postsinaptic este excitat. În alte sinapse are loc o deplasare în sens invers: membrana postsinaptică este hiperpolarizată: valoarea potențialului membranar devine mai mare și devine mai dificilă reducerea acestuia la un nivel critic de depolarizare. Este dificil să excitați o astfel de celulă, este inhibată. Astfel, potenţialul postsinaptic depolarizant este excitator, iar potenţialul hiperpolarizant este inhibitor. În consecință, sinapsele în sine sunt împărțite în excitatorii (care provoacă depolarizarea) și inhibitorii (care provoacă hiperpolarizarea). Sinapsele excitatoare se formează, în cea mai mare parte, pe dendritele celulei postsinaptice, iar pe corpul acesteia se formează sinapsele inhibitorii.
Indiferent de ce se întâmplă pe membrana postsinaptică: depolarizare sau hiperpolarizare, mărimea potențialelor postsinaptice este întotdeauna proporțională cu numărul de molecule transmițătoare care acționează, dar de obicei amplitudinea lor este mică. La fel ca și potențialul receptor, ele se răspândesc de-a lungul membranei pe o distanță foarte scurtă, de exemplu. se referă și la potențialele locale.
Astfel, semnalele de intrare sunt reprezentate de două tipuri de potențiale locale, receptor și postsinaptic, iar aceste potențiale apar în zone strict definite ale neuronului: fie în terminații senzoriale, fie în sinapse. Terminațiile sensibile aparțin neuronilor senzoriali, unde potențialul receptorului ia naștere sub influența oricăror stimuli externi neuronului. Pentru interneuroni, precum și pentru neuronii eferenți, doar potențialul postsinaptic poate fi semnalul de intrare.
3.5. Semnal combinat - potențial de acțiune
Un semnal combinat poate apărea numai într-o regiune a membranei unde există un număr suficient de mare de canale ionice de sodiu. În acest sens, obiectul ideal este dealul axonului - locul în care axonul se îndepărtează de corpul celular, deoarece aici densitatea canalelor pentru sodiu este cea mai mare în întreaga membrană. Astfel de canale sunt dependente de potențial, adică deschis numai atunci când valoarea iniţială a potenţialului de repaus atinge un nivel critic. Potențialul tipic de repaus pentru neuronul mediu este de aproximativ -65 mV, iar nivelul critic de depolarizare corespunde cu aproximativ -55 mV. Prin urmare, dacă este posibilă depolarizarea membranei dealului axonului de la -65 mV la -55 mV, atunci va apărea un potențial de acțiune acolo.
Semnalele de intrare sunt capabile să depolarizeze membrana, de ex. fie potenţiale postsinaptice, fie potenţiale receptor. În cazul potențialelor receptorilor, locul de origine al semnalului combinat este nodul lui Ranvier cel mai apropiat de terminațiile sensibile, unde depolarizarea la un nivel critic este cel mai probabil. În acest sens, trebuie avut în vedere faptul că fiecare neuron senzorial are mai multe terminații, care sunt ramuri ale unui proces. Și, dacă în fiecare dintre aceste terminații, în timpul acțiunii unui stimul, apare un potențial de receptor de amplitudine foarte mică și se răspândește la nodul lui Ranvier cu o scădere a amplitudinii, atunci este doar o mică parte din schimbarea totală de depolarizare. Din fiecare final sensibil, aceste potențiale mici de receptor se deplasează în același timp către cel mai apropiat nod al lui Ranvier, iar în zona de interceptare sunt toate rezumate. Dacă cantitatea totală de depolarizare este suficientă, la interceptare va apărea un potențial de acțiune.
Potențialele postsinaptice care apar pe dendrite sunt la fel de mici ca potențialele receptorului și, de asemenea, scad pe măsură ce se propagă de la sinapsă la dealul axonului, unde poate apărea un potențial de acțiune. În plus, sinapsele hiperpolarizante inhibitorii pot fi în calea propagării potențialelor postsinaptice în întregul corp celular și, prin urmare, posibilitatea depolarizării membranei de deal axonului cu 10 mV pare puțin probabilă. Cu toate acestea, acest rezultat este obținut în mod regulat ca urmare a însumării multor potențiale postsinaptice mici care apar simultan la numeroase sinapse formate de dendritele neuronului cu terminalele axonale ale celulelor presinaptice.
Astfel, semnalul combinat apare, de regulă, ca urmare a însumării a numeroase potențiale locale formate simultan. Această însumare are loc în locul unde există în special multe canale dependente de tensiune și, prin urmare, nivelul critic de depolarizare este mai ușor atins. În cazul integrării potențialelor postsinaptice, un astfel de loc este dealul axonului, iar însumarea potențialelor receptorilor are loc în nodul Ranvier cel mai apropiat de terminațiile senzoriale (sau zona axonului nemielinizat aproape de acestea) . Zona în care apare semnalul combinat se numește integrativ sau trigger (din engleză trigger - trigger).
Termenul englez are succes în expresivitatea sa metaforică, deoarece acumularea de mici schimbări depolarizante se transformă cu viteza fulgerului în zona integrativă într-un potențial de acțiune, care este potențialul electric maxim al celulei și ia naștere în funcție de „totul sau nimic”. principiu. Această regulă trebuie înțeleasă în așa fel încât depolarizarea sub un nivel critic să nu aducă niciun rezultat, iar când se atinge acest nivel, răspunsul maxim este întotdeauna relevat, indiferent de puterea stimulilor: nu există a treia opțiune.
3.6. Conducerea unui potential de actiune
După cum sa menționat deja, amplitudinea semnalelor de intrare este proporțională cu puterea stimulului sau cu cantitatea de neurotransmițător eliberată la sinapsă - astfel de semnale sunt numite graduale. Durata lor este determinată de durata stimulului sau de prezența emițătorului în fanta sinaptică. Amplitudinea și durata potențialului de acțiune nu depind de acești factori: ambii acești parametri sunt în întregime determinați de proprietățile celulei în sine. Prin urmare, orice combinație de semnale de intrare, orice tip de însumare, în condiția unică de depolarizare a membranei la o valoare critică, determină același model standard de potențial de acțiune în zona de declanșare. Are întotdeauna amplitudinea maximă pentru o celulă dată și aproximativ aceeași durată, indiferent de câte ori se repetă condițiile care o cauzează.
După ce a apărut în zona integrativă, potențialul de acțiune se răspândește rapid de-a lungul membranei axonului. Acest lucru se întâmplă din cauza apariției unui curent electric local. Deoarece secțiunea depolarizată a membranei se dovedește a fi încărcată diferit față de cea adiacentă acesteia, între secțiunile încărcate polar ale membranei există o electricitate. Sub influența acestui curent local, zona învecinată este depolarizată la un nivel critic, ceea ce determină apariția unui potențial de acțiune în ea. În cazul unui axon mielinizat, o astfel de secțiune învecinată a membranei este nodul lui Ranvier cel mai apropiat de zona de declanșare, apoi următorul, iar potențialul de acțiune începe să „sare” de la un nod la altul cu o viteză care atinge 100. Domnișoară.
Diferiții neuroni pot diferi unul de celălalt în multe feluri, dar potențialele de acțiune care apar în ei sunt foarte dificil și, în cele mai multe cazuri, imposibil de distins. Aceasta este în cel mai înalt grad un semnal stereotip într-o varietate de celule: senzoriale, interneuroni, motorii. Această stereotipie indică faptul că potențialul de acțiune în sine nu conține nicio informație despre natura stimulului care l-a generat. Puterea stimulului este indicată de frecvența potențialelor de acțiune care apar, iar receptorii specifici și conexiunile interneuronice bine ordonate determină natura stimulului.
Astfel, potențialul de acțiune generat în zona de declanșare se răspândește rapid de-a lungul axonului până la capătul acestuia. Această mișcare este asociată cu formarea de curenți electrici locali, sub influența cărora potențialul de acțiune apare din nou în secțiunea adiacentă a axonului. Parametrii potențialului de acțiune atunci când sunt transportați de-a lungul axonului nu se modifică deloc, ceea ce permite transmiterea informațiilor fără distorsiuni. Dacă axonii mai multor neuroni se găsesc într-un mănunchi comun de fibre, atunci excitația se propagă de-a lungul fiecăruia dintre ei separat.
3.7. Semnal de ieșire
Semnalul de ieșire este adresat unei alte celule sau mai multor celule în același timp și în marea majoritate a cazurilor reprezintă eliberarea unui mesager chimic - un neurotransmițător sau neurotransmițător. În terminațiile presinaptice ale axonului, transmițătorul pre-stocat este stocat în vezicule sinaptice, care se acumulează în zone speciale - zone active. Când potențialul de acțiune atinge terminalul presinaptic, conținutul veziculelor sinaptice este golit în fanta sinaptică prin exocitoză.
Mediatorii chimici ai transmiterii informației pot fi diferite substanțe: molecule mici, precum acetilcolina sau glutamatul, sau molecule peptidice destul de mari - toate acestea sunt sintetizate special în neuron pentru transmiterea semnalului. Odată ajuns în fanta sinaptică, emițătorul difuzează în membrana postsinaptică și se atașează de receptorii săi. Ca urmare a conexiunii receptorilor cu transmițătorul, curentul ionic prin canalele membranei postsinaptice se modifică, iar acest lucru duce la o modificare a valorii potențialului de repaus al celulei postsinaptice, adică. un semnal de intrare apare în el - in în acest caz, potenţial postsinaptic.
Astfel, în aproape fiecare neuron, indiferent de mărimea, forma și poziția sa în lanțul neuronal, pot fi întâlnite 4 zone funcționale: zona receptivă locală, zona integrativă, zona de transmitere a semnalului și zona de ieșire sau secretorie (Fig. 3.3).
În toate organele corpul umanÎn afară de creier, celulele funcționale sunt ținute împreună prin substanțe intercelulare țesut conjunctiv. În sistemul nervos, acest rol este jucat de glia (din greacă glia - lipici), ale cărei celule sunt formate din precursori comuni cu neuronii într-un stadiu incipient al dezvoltării creierului. Glia creează suport pentru neuroni, unește elementele individuale ale sistemului nervos, dar, în același timp, le izolează unele de altele. grupuri diferite neuronii, precum și majoritatea axonilor lor. Astfel, formează structura creierului. Numărul de celule gliale depășește de aproximativ 10 ori numărul de neuroni din creier. Aceste celule diferă unele de altele prin aspect iar în funcţie de funcţia îndeplinită (Fig. 3.4).
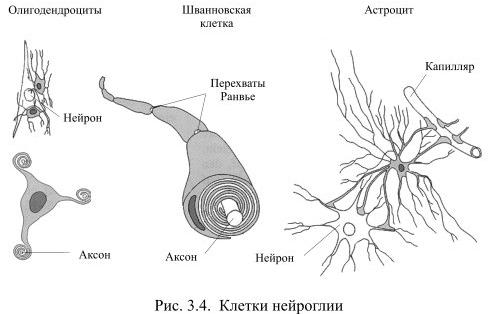
Cele mai frecvente dintre celulele gliale sunt astrocitele, de exemplu, în corpul calos reprezintă 1/4 din toate celulele gliale. Un astrocit are un corp neregulat, în formă de stelat, cu numeroase și relativ lăstari lungi, dintre care unele sunt direcționate către neuroni, iar altele către capilarele sanguine. Aceste procese se extind la capete, formând așa-numitele. picior astrocitar. Pe suprafața capilarului, procesele astrocitelor vecine se închid strâns unele cu altele și învelesc aproape complet vasul de sânge. O astfel de izolare a vasului este una dintre modalitățile de a forma bariera hemato-encefalică - granița dintre sânge și țesut nervos, închis la multe substanțe din sânge.
Alte procese ale astrocitelor învelesc aproape complet corpurile celulare ale neuronilor. Dacă un neuron este excitat pentru o perioadă lungă de timp, concentrația de ioni de potasiu în jurul său crește, iar acest lucru poate reduce excitabilitatea neuronilor vecini. Astrocitele previn această posibilitate prin absorbția excesului de potasiu, acționând astfel ca un tampon. Unele celule gliale devin depolarizate și, deoarece sunt conectate între ele prin joncțiuni interzise, între celulele depolarizate și cele aflate în repaus apare un curent. Acest lucru, cu toate acestea, nu duce la excitare, deoarece există foarte puține canale dependente de tensiune pentru sodiu sau calciu în membrana celulelor gliale. În ciuda faptului că o creștere a concentrației ionilor de potasiu în astrocite modifică unele dintre proprietățile lor, în prezent nu există motive suficiente pentru a le considera participanți direcți la transferul impulsurilor nervoase.
Alte două tipuri de celule gliale: oligodendrocitele și celulele Schwann sunt similare între ele ca aspect și funcție. Ei au corp micși procese relativ mici, aparent aplatizate, care înfășoară în mod repetat axonii neuronilor, oferindu-le astfel o teacă de mielină izolatoare. Mielina este o substanță asemănătoare grăsimii care acționează ca un izolator electric. Atunci când teaca de mielină este pierdută din cauza, de exemplu, a bolilor demielinizante, transmiterea semnalelor de la o parte a creierului la alta este sever perturbată, ducând de obicei la dizabilitate.
Oligodendrocitele creează izolarea cu mielină a axonilor din sistemul nervos central, fiecare oligodendrocit servind de obicei mai mulți axoni. Celulele Schwann învelesc fibrele sistemului nervos periferic cu mielină, fiecare celulă Schwann comitându-se doar cu un axon.
Celulele microgliale sunt împrăștiate în întreaga substanță albă și cenușie a creierului. Spre deosebire de alte celule gliale din creier, acestea sunt străine, extratereștri. Ele sunt formate din monocite din sânge care au reușit să treacă prin pereții capilarelor în creier pentru a se stabili în acesta (în alte țesuturi astfel de monocite sedentare sunt numite macrofage). La fel ca macrofagele altor țesuturi, celulele microgliale acționează ca captatori: ele captează și distrug resturile din celulele în descompunere, această activitate devine deosebit de vizibilă pe fondul leziunilor cerebrale.
Celulele gliale par să joace un rol special în timpul dezvoltării creierului. Unele dintre soiurile lor reglează direcția în care neuronii se deplasează în anumite regiuni ale creierului în creștere, precum și direcția în care cresc axonii. Alte celule gliale pot fi implicate în nutriția celulelor nervoase prin reglarea fluxului sanguin și, prin urmare, transportul de glucoză și oxigen.
rezumat
Într-o varietate remarcabilă caracteristici individuale neuroni individuali sunt descoperite aspecte comune, care fac posibilă clasificarea celulelor nervoase în funcție de structura și funcția lor. Semnalele electrice circulă printr-un neuron într-o singură direcție. În fiecare neuron se pot distinge patru regiuni morfologice care îndeplinesc sarcini funcționale diferite. În fiecare dintre aceste zone, apare un tip special de semnal care este folosit pentru a transmite informații. Celulele gliale, ca și neuronii, diferă în structura și funcția lor.
Întrebări pentru autocontrol
31. Sub literele A-D Sunt indicate diferitele diametre ale axonilor: de-a lungul căruia dintre ele ar trebui să se răspândească mai repede excitația?
A. 0,5 um; B. 1 um; B. 3 um; G. 6 um; D. 9 microni.
32. Cu ce parte a celulei postsinaptice intră cel mai adesea în contact terminalul axonal al celulei presinaptice?
Un corp; B. Soma; V. perikaryon; G. Dendritele; D. Axon.
33. Care dintre următorii neuroni este un neuron eferent?
A. Transmiterea de informații despre durere; B. Transmiterea de informații despre atingerea pielii; B. Transmiterea de informații de la muschii scheletici; D. Transmiterea de informații de la mușchii netezi; D. Transmiterea de informații către glandă.
34. Care regiune morfologică a celulei servește cel mai adesea ca locație a semnalului de intrare?
A. Dendritele; B. Corp; B. Dealul Axon; G. Axon; D. Desinențe axonale.
35. Ce este potențialul receptor?
A. Semnal de intrare; B. Semnal condus; B. Semnal combinat; D. Potenţialul postsinaptic; D. Semnal de ieșire.
36. Ce este un potențial de acțiune?
A. Semnal de intrare; B. Semnal combinat; B. Semnal de ieșire; D. Potențialul local; D. Potenţialul postsinaptic.
37. Ce semnal apare conform regulii „totul sau nimic”?
A. Intrarea; B. Unite; Într-un weekend; G. Postsinaptic; D. Local.
38. Care dintre următoarele semnale este treptat?
A. Potenţial de acţiune; B. Dirijare; B. Postsinaptic; G. Zi liberă; D. Unite.
39. Ce semnal apare în zona de declanșare?
A. Postsinaptic; B. Receptor; B. Intrarea; G. Unite; D. Zi liberă.
40. Care dintre următoarele semnale are cea mai mare amplitudine?
A. Receptor; B. Potenţial de acţiune; B. Postsinaptic; G. Local; D. Intrarea.
41. Ce asigură direct transmisia semnalului de-a lungul axonului?
A. Acţiunea stimulului; B. Eliberarea unui neurotransmițător; B. Prezența acoperirii cu mielină; D. Lipsa acoperirii cu mielina; D. Curentul electric local.
42. Care dintre următoarele este legată de formarea barierei hemato-encefalice?
A. Toate celulele gliale; B. Astrocite; B. Oligodendrocite; G. celule Schwann; D. Microglia.
43. Ce celule asigură izolarea cu mielină a axonilor celulelor sistemului nervos central?
A. Astrocite; B. celule Schwann; B. Oligodendrocite; G. Celule microgliale; D. Toate celulele gliale.
44. Ce funcție îndeplinește mielina?
A. Învelește corpurile neuronilor, oferindu-le protecție mecanică; B. Învelișuri vase de sânge, creând o barieră hemato-encefalică; B. Absoarbe excesul de ioni de potasiu și astfel acționează ca un tampon; D. Este un izolator electric pentru axoni; D. Este conductor de semnale electrice.
45. Când potențialul membranei se deplasează la un nivel critic, ar trebui să apară următoarele:
A. Potenţial de acţiune; B. Potențial receptor; B. Potenţialul postsinaptic; D. Potenţial treptat; D. Semnal de intrare.



