Fájdalom. Fájdalomérzékenység. Nociceptorok. A fájdalomérzékenység útjai. Fájdalomértékelés. A fájdalom kapuja. Opiát peptidek. Fájdalom, gyulladás és egyéb bajok: az érzések másik oldala Fájdalomreceptorok
A fájdalom fiziológiája
A szó szűk értelmében a fájdalom olyan kellemetlen érzés, amely olyan szupererős ingerek hatására jelentkezik, amelyek szerkezeti és funkcionális rendellenességeket okoznak a szervezetben. A fájdalom és az egyéb érzetek között az a különbség, hogy nem tájékoztatja az agyat az inger minőségéről, hanem azt jelzi, hogy az inger káros. A fájdalomérzékelő rendszer másik jellemzője a legösszetettebb és legerősebb efferens vezérlés.
A fájdalomelemző számos programot indít a központi idegrendszerben a szervezet fájdalomra adott válaszára. Ezért a fájdalomnak több összetevője van. A fájdalom szenzoros összetevője kellemetlen, fájdalmas érzésként jellemzi; affektív komponens - erős negatív érzelemként; motivációs komponens – mint negatív biológiai szükséglet, amely kiváltja a szervezet gyógyulásra irányuló viselkedését. A fájdalom motoros komponensét különféle motoros reakciók képviselik: a feltétel nélküli flexiós reflexektől a fájdalomcsillapító viselkedés motoros programjáig. A vegetatív komponens jellemzi a belső szervek diszfunkcióját és az anyagcserét krónikus fájdalom esetén. A kognitív komponens a fájdalom önértékeléséhez kapcsolódik, amelyben a fájdalom szenvedésként hat. Amikor más rendszerek működnek, ezek az összetevők gyengén fejeződnek ki.
A fájdalom biológiai szerepét több tényező határozza meg. A fájdalom a testszövetek fenyegetéséről vagy károsodásáról szól, és figyelmezteti őket. A fájdalomnak kognitív funkciója van: a fájdalom révén az ember megtanulja elkerülni a külső környezet lehetséges veszélyeit. A fájdalom érzelmi összetevője a feltételes reflexek kialakításában a megerősítés funkcióját tölti be. A fájdalom a szervezet védekező és adaptív reakcióit mozgósító tényező, amikor szövetei és szervei károsodnak.
Kétféle fájdalom létezik - szomatikus és zsigeri. A szomatikus fájdalom felületesre és mélyre osztható.
A fájdalomnak három elmélete létezik.
1. Az intenzitáselméletet E. Darwin és A. Goldsteiner javasolta. Ezen elmélet szerint a fájdalom nem specifikus érzés, és nincsenek saját speciális receptorai. Akkor fordul elő, amikor szupererős ingerek hatnak az öt ismert érzékszerv receptoraira. A gerincvelő és az agy impulzusainak konvergenciája és összegzése részt vesz a fájdalom kialakulásában.
2. A specificitás elméletét M. Frey német fiziológus fogalmazta meg. Ezen elmélet szerint a fájdalom egy specifikus érzés, amelynek saját receptor apparátusa, afferens rostjai és a fájdalominformációkat feldolgozó agyi struktúrái vannak. Ez az elmélet később teljesebb kísérleti és klinikai megerősítést kapott.
3. A modern fájdalomelmélet elsősorban a specifikusság elméletén alapul. A specifikus fájdalomreceptorok létezése bizonyított. Ugyanakkor a modern fájdalomelmélet a központi összegzés és a konvergencia fájdalommechanizmusokban betöltött szerepére vonatkozó álláspontot használja. A modern fájdalomelmélet legnagyobb vívmánya a fájdalom központi észlelését és a szervezet fájdalomcsillapító rendszerének beindítását szolgáló mechanizmusok kifejlesztése.
Fájdalom receptorok
A fájdalomreceptorok az érzékeny myelinizált Aδ idegrostok és a nem myelinizált C rostok szabad végződései. Ezek a bőrben, a nyálkahártyákban, a csonthártyában, a fogakban, az izmokban, az ízületekben, a belső szervekben és azok membránjában, valamint az erekben találhatók. Nem találhatók az agy és a gerincvelő idegszövetében. Legnagyobb sűrűségük a dentin és a fogzománc határán található.
A fájdalomreceptorok következő fő típusait különböztetjük meg:
1. Az Aδ-szálak mechanonociceptorai és mechanotermikus nociceptorai erős mechanikai és termikus ingerekre reagálnak, gyors mechanikai és termikus fájdalmat vezetnek, gyorsan alkalmazkodnak; elsősorban a bőrben, az izmokban, az ízületekben, a csonthártyában található; afferens neuronjaik kis befogadó mezőkkel rendelkeznek.
2. A C-rostok poliszenzoros nociceptorai reagálnak mechanikai, termikus és kémiai ingerekre, késői, rosszul lokalizált fájdalmat vezetnek le, és lassan alkalmazkodnak; afferens neuronjaik nagy befogadó mezőkkel rendelkeznek.
A fájdalomreceptorokat háromféle inger stimulálja:
1. Mechanikai irritáló anyagok, amelyek 40 g/mm 2 -nél nagyobb nyomást hoznak létre szorításkor, nyújtáskor, hajlításkor, csavaráskor.
2. A termikus irritáló anyagok lehetnek termikusak (> 45 0 C) és hidegek (< 15 0 С).
3. A sérült szövetsejtekből, hízósejtekből, vérlemezkékből (K +, H +, szerotonin, acetilkolin, hisztamin), vérplazmából (bradikinin, kallidin) és a nociceptív neuronok végződéseiből (P-anyag) felszabaduló kémiai irritáló szerek. Egy részük nociceptorokat (K +, szerotonin, hisztamin, bradikinin, ADP) gerjeszt, mások szenzitizálják.
Fájdalomreceptorok tulajdonságai: a fájdalomreceptoroknak magas a gerjesztési küszöbük, ami csak extrém ingerekre biztosítja a válaszadást. A C-afferensek nociceptorai rosszul alkalmazkodnak a hosszan tartó ingerekhez. Lehetőség van a fájdalomreceptorok érzékenységének növelésére - az irritációjuk küszöbének csökkenése ismételt vagy hosszan tartó stimulációval, amelyet hiperalgéziának neveznek. Ebben az esetben a nociceptorok képesek reagálni a küszöb alatti nagyságrendű ingerekre, valamint más modalitású ingerekre is gerjeszthetők.
Fájdalomérzékenységi utak
Neuronok, amelyek érzékelik a fájdalomimpulzusokat. A törzs, a nyak és a végtagok fájdalomreceptoraiból az első szenzoros neuronok Aδ- és C-rostjai (testük a gerincvelői ganglionokban található) a gerincvelői idegek részeként jutnak el, és a háti gyökereken keresztül bejutnak a gerincvelőbe. , ahol a hátoszlopokban elágazva közvetlenül vagy interneuronokon keresztül szinaptikus kapcsolatokat alakítanak ki a második szenzoros neuronokkal, amelyek hosszú axonjai a spinothalamikus traktusok részét képezik. Ugyanakkor kétféle idegsejtet gerjesztenek: egyes neuronokat csak fájdalmas ingerek aktiválnak, másokat - konvergens idegsejteket - nem fájdalmas ingerek is gerjesztenek. A fájdalomérzékenység második neuronjai túlnyomórészt az oldalsó spinothalamikus traktusok részét képezik, amelyek a fájdalomimpulzusok nagy részét vezetik. A gerincvelő szintjén ezen idegsejtek axonjai az ingerléssel ellentétes oldalra mozdulnak el, az agytörzsben elérik a talamust, és szinapszisokat képeznek a sejtmagok neuronjain. Az első afferens neuronok fájdalomimpulzusainak egy része az interneuronokon keresztül a hajlító izmok motoros neuronjaira kapcsol át, és részt vesz a védő fájdalomreflexek kialakításában. A laterális spinothalamikus traktusban az evolúciósan fiatalabb neospinothalamikus traktus és az ősi paleospinothalamikus traktus különböztethető meg.
A neospinothalamikus pálya az Aδ rostok mentén főleg a thalamus specifikus szenzoros (ventral posterior) magjaihoz vezeti a fájdalomjeleket, amelyek jó topográfiai vetülettel rendelkeznek a test perifériájára. Ezenkívül az impulzusok egy kis része a törzs retikuláris képződményébe, majd a talamusz nem specifikus magjaiba kerül. Ennek az útvonalnak a szinapszisaiban a gerjesztés átvitele a gyorsan ható glutamát transzmitter segítségével történik. A thalamus meghatározott magjaiból a fájdalomjelek túlnyomórészt az agyféltekék szenzoros kéregébe kerülnek. Ezek a tulajdonságok alkotják a neospinothalamikus út fő funkcióját - a „gyors” fájdalom levezetését és annak nagyfokú lokalizációval történő érzékelését.
A paleospinothalamikus pálya elsősorban a thalamus nem specifikus magjaihoz vezet fájdalomjeleket a C-szálak mentén közvetlenül vagy az agytörzs retikuláris formációjának neuronjaiban történő átkapcsolás után. A gerjesztés átvitele a szinapszisokon ezen az úton lassabban megy végbe. A közvetítő a P anyag. A nem specifikus magokból impulzusok jutnak az agykéreg érzékszervi és egyéb részeibe. Az impulzus egy kis része a talamusz meghatározott magjaiba is bejut. Alapvetően ennek az útvonalnak a rostjai a thalamus nem specifikus magjainak neuronjain végződnek; 2) retikuláris képződés; 3) központi szürkeállomány; 4) kék folt; 5) hipotalamusz. A „késői”, rosszul lokalizált fájdalom a paleospinothalamikus úton továbbítódik, és kialakulnak a fájdalomérzékenység affektív és motivációs megnyilvánulásai.
Ezenkívül a fájdalomérzékenység részben más felszálló pályákon keresztül történik: az elülső spinothalamikus, vékony és ék alakú pályákon.
A fenti utak más típusú érzékenységet is vezetnek: a hőmérsékletet és a tapintást.
Az agykéreg szerepe a fájdalomérzékelésben
A fájdalom teljes szenzoros érzékelése a test által az agykéreg részvétele nélkül lehetetlen.
A fájdalomanalizátor elsődleges vetületi tere a hátsó központi gyrus szomatoszenzoros kéregében található. Lehetővé teszi a „gyors” fájdalom érzékelését és a testen belüli elhelyezkedésének azonosítását. A fájdalom helyének pontosabb azonosítása érdekében a folyamat szükségszerűen magában foglalja az elülső központi gyrus motoros kéregének neuronjait.
A másodlagos vetületi mező a szomatoszenzoros kéregben található, a centrális sulcus és a temporális lebeny felső szélének metszéspontjában. Ennek a mezőnek a neuronjai kétoldalú kapcsolatban állnak a thalamus magjaival, ami lehetővé teszi, hogy ez a mező szelektíven szűrje a thalamuson áthaladó fájdalmas izgalomokat. Ez pedig lehetővé teszi, hogy ez a terület részt vegyen a szükséges viselkedési aktus engramjának emlékezetből való előhívásával, az effektorok tevékenységében történő megvalósításával és az elért hasznos eredmény minőségének felmérésével kapcsolatos folyamatokban. A fájdalomviselkedés motoros összetevői a motoros és premotoros kéreg, a bazális ganglionok és a kisagy együttes tevékenységében jönnek létre.
A frontális kéreg fontos szerepet játszik a fájdalomérzékelésben. A fájdalom (kognitív összetevője) önértékelését és a célzott fájdalomviselkedés kialakítását biztosítja.
A limbikus rendszer (gyrus cingulate, hippocampus, gyrus fogazat, a halántéklebeny amygdala komplexuma) a thalamus elülső magjaiból kapja a fájdalominformációkat és a fájdalom érzelmi összetevőjét képezi, vegetatív, szomatikus és viselkedési reakciókat vált ki, amelyek adaptív reakciókat biztosítanak a thalamushoz. fájdalmas inger.
A fájdalom bizonyos fajtái
Vannak fájdalmak, amelyeket elneveznek kivetítés vagy fantom. Előfordulásuk a fájdalom vetületének törvényén alapul: függetlenül attól, hogy az afferens pálya melyik része irritált, a fájdalom ezen érzékszervi útvonal receptorainak területén érezhető. A modern adatok szerint a fájdalomérzékelési rendszer minden része részt vesz az ilyen típusú fájdalom kialakulásában.
Vannak még ún tükröződött fájdalom: amikor a fájdalom nemcsak az érintett szervben, hanem a test megfelelő dermatómájában is érezhető. A megfelelő dermatóma testfelületének azon területeit, ahol a fájdalomérzet jelentkezik, nevezzük Zakharyin–Ged zónák. A hivatkozott fájdalom előfordulása annak köszönhető, hogy az érintett szerv receptoraiból fájdalomimpulzusokat hordozó neuronok és a megfelelő dermatóma bőre a spinothalamikus traktus ugyanazon neuronján konvergálnak. Ennek a neuronnak az érintett szerv receptoraitól való irritációja a fájdalom vetületének törvényével összhangban ahhoz a tényhez vezet, hogy a fájdalom a bőrreceptorok területén is érezhető.
Antinociceptív rendszer
A fájdalomcsillapító rendszer négy szintből áll: gerincvelő, agytörzs, hipotalamusz és kortikális.
1. Az antinociceptív rendszer spinális szintje. Fontos eleme a gerincvelő „kapuszabályozása”, melynek koncepciója a következő alapelveket tartalmazza: a fájdalom idegimpulzusainak átvitele az első idegsejtektől a spinothalamikus traktus neuronjaihoz (második neuronok) a gerincvelő hátsó oszlopaiban. a gerincvelőt a gerinckapu-mechanizmus modulálja - a gerincvelő zselatinos anyagában található gátló neuronok. A különböző szenzoros pályák elágazó axonjai ezeken az idegsejteken végződnek. A zselatinos anyag neuronjai viszont preszinaptikus gátlást fejtenek ki a fájdalom első és második idegsejtjének átkapcsolási helyén és más szenzoros pályákon. Egyes neuronok konvergensek: a neuronok nem csak a fájdalomreceptorokból, hanem más receptorokból is szinapszisokat képeznek rajtuk. A gerincportál szabályozását a nagy átmérőjű (nem fájdalomérzékenység) és a kis átmérőjű (fájdalomérzékenység) afferens rostok mentén érkező impulzusok aránya szabályozza. Az intenzív impulzusáramlás a nagy átmérőjű rostok mentén korlátozza a fájdalomjelek átvitelét a spinothalamikus pályák idegsejtjei felé (bezárja a „kaput”). Éppen ellenkezőleg, a fájdalomimpulzusok intenzív áramlása az első afferens neuron mentén, gátolva a gátló interneuronokat, megkönnyíti a fájdalomjelek továbbítását a spinothalamikus pályák neuronjaihoz (kinyitja a „kaput”). A gerinckapu mechanizmusa az agytörzsi struktúrákból származó idegimpulzusok állandó befolyása alatt áll, amelyek leszálló pályákon továbbítják mind a substantia gelatinosa, mind a spinothalamikus traktus neuronjaihoz.
2. Az antinociceptív rendszer agytörzsi szintje. A fájdalomcsillapító rendszer törzsszerkezetei közé tartozik egyrészt a központi szürkeállomány és a raphe sejtmagok, amelyek egyetlen funkcionális blokkot alkotnak, másrészt a retikuláris formáció magnocelluláris és paragiáns sejtmagjai, valamint a locus coeruleus. Első komplexus blokkolja a fájdalomimpulzusok áthaladását a gerincvelő hátsó szarvának magjainak közvetítő neuronjainak, valamint a trigeminus ideg szenzoros magjainak közvetítő neuronjainak szintjén, kialakítva a fájdalomérzékenység felszálló pályáit. A második komplex szinte az egész antinociceptív rendszert gerjeszti (lásd 1. ábra).
3. Az antinociceptív rendszer hipotalamusz szintje egyrészt önállóan működik, másrészt az antinociceptív mechanizmusokat törzsszinten irányító és szabályozó környezetként működik a különböző nukleáris hovatartozású hipotalamusz neuronjai és a különböző neurokémiai sejtek közötti kapcsolatok miatt. sajátosságait. Közülük neuronokat azonosítottak, amelyek végződéseiben enkefalin, β-endorfin, noradrenalin és dopamin szabadul fel (lásd 2. ábra).
4. Az antinociceptív rendszer corticalis szintje. Az agykéreg szomatoszenzoros területe egyesíti és szabályozza az antinociceptív struktúrák aktivitását különböző szinteken. Ebben az esetben a legfontosabb szerep az aktiválásban gerinc- és szárszerkezetek a másodlagos szenzoros területet játssza. Neuronjai alkotják a fájdalomérzékenység csökkenő szabályozásának legnagyobb számú rostját, amelyek a gerincvelő hátsó szarvaihoz és az agytörzs magjaihoz vezetnek. A másodlagos szenzoros kéreg módosítja az antinociceptív rendszer szárkomplexének aktivitását. Ezenkívül az agykéreg szomatoszenzoros mezői szabályozzák az afferens fájdalomimpulzusok átvezetését a thalamuson keresztül. A thalamuson kívül az agykéreg szabályozza a fájdalomimpulzusok áthaladását a hipotalamuszban, a limbikus rendszerben, a retikuláris képződésben és a gerincvelőben. A cortico-hypothalamus hatások biztosításában a vezető szerepet a frontális kéreg neuronjai jelölik.
Az antinociceptív rendszer közvetítői
A fájdalomcsillapító rendszer közvetítői közé tartoznak az agyban, az adenohipofízisben, a mellékvese velőben, a gyomor-bél traktusban, az inaktív prekurzorokból származó placentában képződő peptidek. Az antinociceptív rendszer opiát mediátorai közé tartoznak: 1) ά-, β-, γ-endorfinek; 2) enkefalinok; 3) dinorfinok. Ezek a mediátorok háromféle opiátreceptorra hatnak: μ-, δ-, κ-receptorokra. A μ-receptorok legszelektívebb stimulátorai az endorfinok, a δ-receptorok az enkefalinok, a κ-receptorok a dinorfinok. A μ- és κ-receptorok sűrűsége magas az agykéregben és a gerincvelőben, az agytörzsben átlagos; a δ-receptorok sűrűsége az agykéregben és a gerincvelőben átlagos, az agytörzsben alacsony. Az opioid peptidek gátolják a fájdalmat okozó anyagok hatását a nociceptorok szintjén, csökkentik a fájdalomimpulzusok ingerlékenységét és vezetőképességét, valamint gátolják a fájdalomimpulzusokat továbbító körökben elhelyezkedő neuronok kiváltott reakcióját. Ezek a peptidek vérrel és agy-gerincvelői folyadékkal jutnak el a fájdalomérző rendszer neuronjaihoz. Az opioid mediátorok a fájdalomcsillapító rendszer neuronjainak szinaptikus végződéseiben szabadulnak fel. Az endorfinok fájdalomcsillapító hatása az agyban és a gerincvelőben magas, az enkefalinek hatása ezekben a struktúrákban közepes, a dinorfin hatása az agyban alacsony, a gerincvelőben pedig erős.
1. ábra. Az első szintű fájdalomcsillapító rendszer fő elemeinek kölcsönhatása: agytörzs - gerincvelő. (a nyitott körök serkentő neuronok, a fekete körök gátlók).
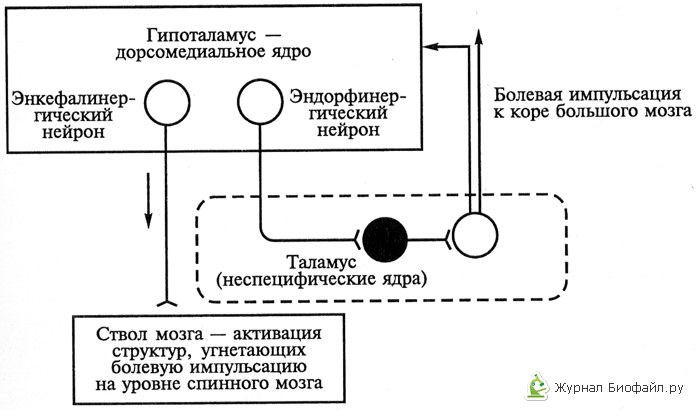
2. ábra. A szervezet másodfokú fájdalomcsillapító rendszerének (hipotalamusz - thalamus - agytörzs) működési mechanizmusa opioidok felhasználásával.
A világos körök serkentő neuronok, a fekete körök gátló neuronok.
A fájdalom súlyosságát nem önmagában az exogén vagy endogén fájdalom erőssége határozza meg. Ez nagyban függ a fájdalomrendszer nociceptív és antinociceptív részének aktivitásának arányától, aminek adaptív jelentősége van.
A "Hőmérsékletérzékenység. Visceralis érzékenység. Vizuális érzékszervi rendszer" témakör tartalomjegyzéke:1. Hőmérsékletérzékenység. Hőreceptorok. Hideg receptorok. Hőmérséklet érzékelése.
2. Fájdalom. Fájdalomérzékenység. Nociceptorok. A fájdalomérzékenység útjai. Fájdalomértékelés. A fájdalom kapuja. Opiát peptidek.
3. Visceralis érzékenység. Visceroreceptorok. Visceralis mechanoreceptorok. Visceralis kemoreceptorok. Visceralis fájdalom.
4. Vizuális szenzoros rendszer. Vizuális észlelés. Fénysugarak vetítése a szem retinájára. A szem optikai rendszere. Fénytörés.
5. Szállás. A tiszta látás legközelebbi pontja. Szálláskínálat. Távollátás. Életkorral összefüggő távollátás.
6. Fénytörési hibák. Emmetropia. Rövidlátás (myopia). Távollátás (hipermetrópia). Asztigmatizmus.
7. Pupilláris reflex. A látómező vetítése a retinára. Binokuláris látás. A szemek konvergenciája. A szemek eltérése. Keresztirányú eltérés. Retinotópia.
8. Szemmozgások. Szemmozgások követése. Gyors szemmozgások. Központi mélyedés. Saccades.
9. Fényenergia átalakítása a retinában. A retina funkciói (feladatai). Vakfolt.
10. Scotopikus retinarendszer (éjszakai látás). A retina fotopikus rendszere (nappali látás). A retina kúpjai és rudai. Rhodopszin.
Fájdalom. Fájdalomérzékenység. Nociceptorok. A fájdalomérzékenység útjai. Fájdalomértékelés. A fájdalom kapuja. Opiát peptidek.
Fájdalom tényleges vagy potenciális szövetkárosodáshoz kapcsolódó kellemetlen szenzoros és érzelmi élményként definiálják, vagy ilyen károsodással írják le. Más érzékszervi módoktól eltérően a fájdalom szubjektíven mindig kellemetlen, és nem annyira információforrásként szolgál a környező világról, hanem károsodás vagy betegség jelzéseként. Fájdalomérzékenységösztönzi a káros környezeti tényezőkkel való érintkezés megszüntetését.
Fájdalom receptorok vagy nociceptorok szabad idegvégződések a bőrben, a nyálkahártyákban, az izmokban, az ízületekben, a csonthártyában és a belső szervekben. Az érzékeny végződések vagy nem myelinizált, vagy vékony myelinizált rostokhoz tartoznak, amelyek meghatározzák a jelátvitel sebességét a központi idegrendszerben, és különbséget tesznek a korai, rövid és akut fájdalom között, amely akkor jelentkezik, amikor az impulzusokat nagyobb sebességgel hajtják végre. a myelinizált rostok mentén, valamint késői, tompa és hosszan tartó fájdalom, nem pulpa rostok mentén történő jelátvitel esetén. Nociceptorok multimodális receptorokhoz tartoznak, mivel különböző természetű ingerekre aktiválhatók: mechanikai (ütés, vágás, szúrás, csípés), termikus (meleg vagy hideg tárgyak hatása), kémiai (hidrogénionok koncentrációjának változása, hisztamin, bradikinin és számos más biológiailag aktív anyag). Nociceptor érzékenységi küszöb magas, tehát csak a kellően erős ingerek okozzák a primer szenzoros neuronok gerjesztését: például a mechanikai ingerek fájdalomérzékenységi küszöbe megközelítőleg ezerszer magasabb, mint a tapintási érzékenység küszöbe.
Az elsődleges szenzoros neuronok központi folyamatai a háti gyökerek részeként belépnek a gerincvelőbe, és szinapszisokat képeznek a gerincvelő hátsó szarvaiban található másodrendű neuronokkal. A másodrendű neuronok axonjai a gerincvelő ellenkező oldalára mozognak, ahol a spinothalamikus és a spinoretikuláris traktusokat alkotják. Spinothalamikus traktus a thalamus inferior posterolaterális magjának neuronjain végződik, ahol a fájdalom és a tapintási érzékenység útvonalainak konvergenciája következik be. A thalamus idegsejtjei a szomatoszenzoros kéregre vetítést alkotnak: ez az út biztosítja a fájdalom tudatos érzékelését, lehetővé teszi az inger intenzitásának és lokalizációjának meghatározását.
Rostok spinoretikuláris traktus A retikuláris formáció neuronjain végződnek, amelyek kölcsönhatásba lépnek a talamusz mediális magjaival. A fájdalmas stimuláció során a talamusz mediális magjainak neuronjai moduláló hatást fejtenek ki a kéreg nagy területeire és a limbikus rendszer struktúráira, ami az emberi viselkedési aktivitás növekedéséhez vezet, és érzelmi és autonóm reakciók kísérik. Ha a spinothalamikus pálya a fájdalom szenzoros tulajdonságainak meghatározására szolgál, akkor a spinoretikuláris pálya általános riasztójelként szolgál, és általános stimuláló hatást fejt ki az emberre.

Szubjektív fájdalomértékelés meghatározza mindkét út idegi aktivitásának arányát és az attól függő antinociceptív leszálló pályák aktiválódását, ami megváltoztathatja a jelek vezetési jellegét nociceptorok. Az érzékszervi rendszerhez fájdalomérzékenység egy endogén mechanizmus ennek csökkentésére a gerincvelő hátsó szarvaiban a szinaptikus átkapcsolások küszöbének szabályozásával (“ a fájdalom kapuja"). A gerjesztés átvitelét ezeknél a szinapszisoknál befolyásolják a szürkeállomány neuronjainak leszálló rostjai a vízvezeték, a locus coeruleus és a median raphe egyes magjai körül. Ezen neuronok mediátorai (enkefalin, szerotonin, noradrenalin) gátolják a másodrendű neuronok aktivitását a gerincvelő hátsó szarvában, ezáltal csökkentve a nociceptorokból érkező afferens jelek vezetését.
Fájdalomcsillapító (fájdalomcsillapítók) hatással vannak opiát peptidek (dinorfin, endorfinok), amelyeket a hipotalamusz neuronjai szintetizálnak, amelyek hosszú folyamatokkal behatolnak az agy más részeibe. Opiát peptidek a limbikus rendszer és a thalamus mediális régiójának neuronjainak specifikus receptoraihoz kötődnek, kialakulásuk fokozódik bizonyos érzelmi állapotok, stressz, hosszan tartó fizikai aktivitás hatására, terhes nőknél röviddel a szülés előtt, valamint pszichoterápiás hatások, ill. akupunktúra. A fokozott oktatás eredményeként opiát peptidek Az antinociceptív mechanizmusok aktiválódnak, és a fájdalomérzékenységi küszöb nő. A fájdalomérzet és annak szubjektív megítélése közötti egyensúly a fájdalmas ingerek észlelésének folyamatában részt vevő elülső agyterületek segítségével jön létre. Ha a homloklebenyek sérültek (például sérülés vagy daganat miatt) fájdalomküszöb nem változik, így a fájdalomérzékelés szenzoros komponense változatlan marad, azonban a fájdalom szubjektív érzelmi megítélése más lesz: csak szenzoros érzetként kezdik felfogni, nem szenvedésként.
Leírják az izomfájdalom-receptorok aktiválódásához vezető tényezőket is: mechanikai trauma, az erek és az izomrostok integritásának megzavarása, a hidrogénionok fokozott koncentrációja.
Izomfájdalom receptorok (nociceptorok)
A nociceptív fájdalom fogalma
A fájdalom egy speciális érzékenység, amely egy patogén inger hatásához kapcsolódik, és szubjektíven kellemetlen érzésekkel jellemezhető. A fájdalmat a szervezetben végbemenő jelentős változások is jellemzik, beleértve a létfontosságú funkcióinak súlyos megzavarását és akár a halált is.
Nociceptív nevezzük a fájdalmat, amelyet bármely tényező (mechanikai trauma, égés, gyulladás stb.) a perifériás fájdalomreceptorokra gyakorolt hatása okoz, az idegrendszer más részeinek károsodása nélkül.
Érzékszervi idegek és receptorok
A fájdalomérzékenységért az Aδ típusú és a C-szálú érzékrostok felelősek. Ezeket a rostokat csak nagyon erős fájdalmas stimuláció gerjeszti. Amikor blokkolják őket, a fájdalomérzékenység teljesen eltűnik. Az Aδ és C rostok végei fájdalomreceptorok. Ezek a rostok beidegzik a bőrt, a mély szöveteket, a belső szerveket és az izmokat.
Az izomfájdalom-receptorok (nociceptorok) elhelyezkedése
A fájdalmas idegvégződések egyenetlenül oszlanak el a testben. Hálózatként borítják az egész bőrt. Kisebb mennyiségben vannak jelen az izmokban. Az izomfájdalom-receptorok diffúzan helyezkednek el az izomrostok között, az izomrostokat körülvevő kötőszöveti membránokban és az izom egészében, valamint az izom-tendinos csomópont területén. Fájdalomimpulzusokat vezetnek az A mentén lévő izomból δ -rostok és C-rostok az agykéregbe, ahol a nociceptorokból származó impulzusaktivitás növekedését fájdalomérzetként érzékelik.
Az izomnociceptorok aktiválása
Az izomnociceptorokat az intenzív károsító mechanikai hatások könnyen gerjesztik. Az izomrostok között és az ínben elhelyezkedő fájdalomreceptorok aktiválódását és fokozott érzékenységét számos patofiziológiai állapot okozhatja. A legismertebb lehetőség az akut trauma.
Az izomfájdalom-receptorok aktiválódását nemcsak mechanikusan, hanem az erek és az izomrostok integritásának megsértése is okozhatja. Ennek eredményeként megnő az endogén anyagok koncentrációja a szövetben, ami a nociceptorok érzékenységének növekedését okozza. A fájdalmat okozó anyagok közé tartozik a hidrogénionok (H +) nagy koncentrációja. Ismeretes, hogy az izomrostok hipertrófiáját célzó erőgyakorlatok végzése során a laktát felhalmozódik bennük, és nő a hidrogénionok koncentrációja. Ez az egyik oka annak, hogy fájdalmat okoz az izmokban.
Irodalom
- Alekseev V.V. Myogenic fájdalom szindrómák: patogenezis és terápia // Hatékony gyógyszeres terápia, 2011.- T. 17. P. 30-34.
- Boxer O.Ya., Grigoriev K.I. A fájdalom tudománya: kórélettani és orvosi-pszichológiai vonatkozások // Nővér, 2005.- 8.- P.2-5.
Fájdalmas irritációk léphetnek fel a bőrön, a mély szövetekben és a belső szervekben. Ezeket az ingereket a test egészében elhelyezkedő nociceptorok érzékelik, kivéve az agyat. A mikroneurográfiai technika lehetővé tette annak megállapítását, hogy az embernek ugyanaz a kétféle fájdalomreceptorja (nociceptora) van, mint a többi emlősnek. Anatómiailag a nociceptorok első típusát szabad idegvégződések képviselik, amelyek fa formájúak (mielinrostok). Ezek gyors A-delta rostok, amelyek 6-30 ms sebességgel hajtanak végre stimulációt. Ezeket a szálakat a bőr nagy intenzitású mechanikai (tűszúrás) és néha termikus irritációi gerjesztik. Az A-delta nociceptorok elsősorban a bőrben találhatók, beleértve az emésztőrendszer mindkét végét. Az ízületekben is megtalálhatók. Az A-delta szálak adója ismeretlen.
A nociceptorok másik típusát a sűrű, nem kapszulázott glomeruláris testek képviselik (nem myelinizált C rostok, amelyek 0,5-2 ms sebességgel stimulálják). Ezeket az afferens rostokat az emberben és más főemlősökben polimodális nociceptorok képviselik, és ezért reagálnak a mechanikai, hőmérsékleti és kémiai stimulációra egyaránt. A szövetkárosodás során keletkező vegyi anyagok aktiválják őket, egyben kemoreceptorok is, és evolúciós primitívségükkel optimális szövetkárosító receptornak számítanak. A C-rostok a központi idegrendszer kivételével minden szövetben eloszlanak. A perifériás idegekben azonban nervi nervorum néven vannak jelen. A szövetkárosodást érzékelõ receptorokkal rendelkezõ rostok P anyagot tartalmaznak, amely transzmitterként mûködik. Ez a típusú nociceptor tartalmazza a kalcitonin génnel rokon peptidet, valamint a belső szervekből származó rostokat - vazoaktív intestinalis peptidet (Nicholls és mtsai, 1992).
A gerincvelő hátsó szarvai
A legtöbb „fájdalomrost” a gerincvelői idegeken keresztül jut el a gerincvelőbe (ha a nyakból, a törzsből és a végtagokból erednek), vagy a háromosztatú ideg részeként a medulla oblongatába jut. A dorsalis gyökér ganglionhoz közel, a gerincvelőbe való belépés előtt a hátgyökér egy mediális részre oszlik, amely vastag mielinrostokat tartalmaz, és egy oldalsó részre, amely vékony mielin (A - delta) és nem myelin (C) rostokat tartalmaz. (Sindou, et al., 1975), amely lehetővé teszi a sebész számára, hogy működési mikroszkóp segítségével elvégezze funkcionális szétválasztásukat. Ismeretes azonban, hogy a C rostok hozzávetőleg 30%-ának proximális axonjai, miután elhagyták a gangliont a gerincvelőből, visszatérnek a szenzoros és motoros gyökerek (zsinór) ízületi pályájába, és az elülső gyökereken keresztül belépnek a gerincvelőbe (Coggeshall). et al., 1975). Valószínűleg ez a jelenség magyarázza a fájdalom csillapítására irányuló dorzális rhizotómia kudarcát (Blumenkopf, 1994). Mindazonáltal, mivel minden C-rost neuronjait a dorsalis ganglionba helyezi, a cél gangliolízissel elérhető (Nash, 19986). Amikor a nociceptív rostok belépnek a gerincvelőbe, felszálló és leszálló ágakra osztódnak. Mielőtt a hátsó szarv szürkeállományában véget érnének, ezek a rostok a gerincvelő több szegmensére irányíthatók. Elágazással kapcsolatot alakítanak ki számos más idegsejttel. Így a „posthorn komplexum” kifejezés erre a neuroanatómiai struktúrára utal. A poszthorn-relésejtek két fő osztályát közvetlenül vagy közvetve aktiválja a nociceptív információ: a „nociceptív specifikus” neuronokat, amelyeket csak nociceptív ingerek aktiválnak, és a „széles dinamikus tartományú” vagy „konvergens” neuronokat, amelyeket szintén nem nociceptív ingerek aktiválnak. A gerincvelő háti szarvának szintjén nagyszámú elsődleges afferens inger közvetítődik interneuronokon vagy asszociatív neuronokon keresztül, amelyek szinapszisai elősegítik vagy megakadályozzák az impulzusok átvitelét. A perifériás és központi vezérlés a sejtréteg melletti kocsonyás anyagban található.
A kapuvezérlés, mint belső gerincmechanizmus.
A „kapukontroll” elmélete a fájdalommechanizmusok egyik legtermékenyebb koncepciója (Melzack, Wall, 1965), bár anatómiai és fiziológiai alapja még mindig nem teljesen kidolgozott (Swerdlow, Charlton, 1989). Az elmélet fő álláspontja az, hogy a vékony ("fájdalom") perifériás rostokon áthaladó impulzusok megnyitják az idegrendszer "kapuját", hogy elérjék annak központi részeit. Két körülmény zárhatja be a kaput: a vastag („tapintható”) rostokon áthaladó impulzusok és bizonyos, az idegrendszer magasabb részeiből leszálló impulzusok. A kaput bezáró vastag perifériás rostok hatásmechanizmusa az, hogy a mély szövetekben, például izmokban és ízületekben fellépő fájdalmat csökkenti az irritáció ellen - a bőrfelület mechanikus dörzsölése vagy irritáló kenőcsök használata (Barr, Kiernan, 1988). ). Ezeknek a tulajdonságoknak terápiás alkalmazásai vannak, mint például a vastag bőrrostok magas frekvenciájú, alacsony intenzitású elektromos stimulációja (Wall és Sweet, 1967), amely transzkután elektromos idegstimuláció (TENS) néven ismert, vagy vibrációs stimuláció (Lunderberg, 1983). . A második mechanizmus (a kapu belülről történő bezárása) akkor lép működésbe, amikor az agytörzsből leszálló gátló rostok aktiválódnak, akár közvetlen stimulációval, akár heteroszegmentális akupunktúrával (alacsony frekvenciájú, nagy intenzitású perifériás stimuláció). Ebben az esetben a leszálló rostok aktiválják a háti szarvak felületes rétegeiben elhelyezkedő interneuronokat, amelyek posztszinaptikusan gátolják a kocsonyás sejteket, ezáltal megakadályozzák az információ magasabbra továbbítását (Swerdlow, Charlton, 1989).
Opioid receptorok és mechanizmusok.
Az opioid peptidek és opioid receptorok felfedezése a 70-es évek elejére nyúlik vissza. 1973-ban három kutatócsoport (Hughes, Kosterlitz, Yaksh) azonosította a morfium alkalmazási helyeit, két évvel később pedig két másik csoport fedezte fel a morfium hatását utánzó természetes peptidek lokalizációját. Az opioid receptorok három osztálya klinikai jelentőséggel bír: mukappa és delta receptorok (Kosterlitz, Paterson, 1985). Eloszlásuk a központi idegrendszeren belül nagyon változó. A receptorok sűrű eloszlása a gerincvelő hátsó szarvaiban, a középagyban és a talamuszban található. Az immuncitokémiai vizsgálatok a gerincvelő hátsó szarvának felszíni rétegeiben mutatták ki a spinális opioid receptorok legmagasabb koncentrációját. Az endogén opioid peptidek (enkefalin, endorfin, dinorfin) kölcsönhatásba lépnek az opioid receptorokkal, amikor fájdalmas ingerek lépnek fel a fájdalomküszöb leküzdése következtében. Az a tény, hogy sok opioid receptor található a gerincvelő felszíni rétegeiben, azt jelenti, hogy az opiátok könnyen bejuthatnak a környező agy-gerincvelői folyadékból. Az opiátok közvetlen spinális hatásának kísérleti megfigyelései (Yaksh, Rudy, 1976) lehetővé tették terápiás alkalmazásukat intratekális (Wang, 1977) és epidurális (Bromage és mtsai, 1980) beadásával.
Ismeretes, hogy nagy dózisú morfiumra van szükség a gerincvelői neuronok túlzott ingerlékenységének visszaszorításához. Ha azonban kis dózisú morfiumot közvetlenül a károsító stimuláció előtt írnak fel, akkor a kiváltó központi hiperingerlékenység soha nem alakul ki (Woolf, Wall, 1986). Mára világossá vált, hogy az előkezelés megelőzheti a súlyos posztoperatív fájdalmakat (Wall, Melzack, 1994).
A fájdalom emelkedő ösvényei.
Régóta ismert, hogy a felszálló „fájdalomutak” a gerincvelő fehérállományának anterolaterális zsinórjaiban helyezkednek el, és a fájdalmas ingerek belépési oldalával ellentétes oldalon futnak (Spiller, 1905). Az is jól ismert, hogy a spinothalamikus és a spinoreticularis pálya rostjainak egy része, amelyek fájdalomstimulációt hajtanak végre, jelen vannak a posterolaterális zsinórban (Barr, Kiernan, 1988). a spinothalamikus és a spinoretikuláris traktusok szinte teljes elvesztéséhez vezet a fájdalomérzet a test másik oldalán a sérülés szintje alatt (Kaye, 1991). Általában azonban az érzékenység fokozatosan, néhány hét alatt helyreáll, ami a szinaptikus átszervezéssel és az érintetlen alternatív utak toborzásával magyarázható. A commissural myelotomia elhúzódó fájdalomcsillapítást okoz az érintett szegmensekben.
A spinothalamikus traktus két részre osztható:
- 1. Neospinothalamikus traktus (gyors vezetés, monoszinaptikus transzmisszió, jól lokalizált (epikritikus) fájdalom, A - rostok). Ez a traktus a thalamus specifikus laterális magjaihoz (ventroposterolaterális és ventroposteromedialis magokhoz) megy.
- 2. Paleospinothalamikus rendszer (poliszinaptikus transzmisszió, lassú vezetés, rosszul lokalizált (protopathiás) fájdalom, C rostok). Ezek az utak a nem specifikus mediális thalamus magokhoz (mediális mag, intralamináris mag, median center) emelkednek. A thalamus mediális magjai felé haladva a traktus néhány rostot küld a retikuláris formációba.
A talamuszban elhelyezett sztereotaktikus elektródák lehetővé teszik ezen struktúrák sajátos patofiziológiájának felismerését, és a thalamus mediális (főleg nucl. centralis lateralis) és laterális (nucl. ventroposterior) magjai közötti egyensúly meglétén alapuló koncepció kidolgozását. melynek megsértése mindkettőjük túlzott gátlásához vezet a reticularis thalamus mag által, majd a fájdalommal járó kérgi mezők paradox aktiválásához. A mediális sztereotaxiás thalamotomia újrakezdése, figyelembe véve az új technikai, anatómiai és fiziológiai adatokat, a krónikus és kezelésre rezisztens perifériás és centrális neurogén fájdalomban szenvedő betegek kétharmadának 50-100%-kal enyhül (Jeanmonod et al., 1994).
A neospinothalamikus rendszeren keresztül bejutó impulzusok olyan rostokra kapcsolódnak át, amelyek a belső kapszula hátsó combján keresztül a kéreg első szomatoszenzoros területére, a posztcentrális gyrusra és a második szomatoszenzoros területre (operculum parietal) továbbítják a jeleket. A thalamus laterális magjában a lokális szerveződés magas foka lehetővé teszi a fájdalom térbeli lokalizációját. Mindkét világháborúban több ezer agykérgi elváltozást vizsgáltak, és azt mutatják, hogy a posztcentrális gyrus sérülése soha nem okoz fájdalomérzékenységet, bár a szomatotopikusan szervezett, alacsony küszöbű mechanoreceptív érzet elvesztéséhez, valamint tűszúrás érzéséhez (Bowsher) , 1987).
A paleospinothalamikus traktuson át bejutó impulzusok a thalamus medialis magjába kapcsolódnak, és diffúz módon a neocortexbe vetülnek. A frontális régióban lévő projekció a fájdalom affektív összetevőit tükrözi. A pozitronemissziós tomográfia azt mutatja, hogy a káros ingerek aktiválják a neuronokat a gyrus cingulárisban és az orbitális frontális kéregben (Jones és mtsai, 1991). A cingulotómia vagy prefrontális lobotómia kiváló eredményeket mutatott a rákos betegek fájdalmának kezelésében (Freeman, Watts, 1946). Így az agyban nincs „fájdalomközpont”, a fájdalom észlelése és a fájdalomra adott válasz a központi idegrendszer egészének függvénye (Diamond, Coniam, 1991, Talbot és mtsai, 1991).
Csökkenő fájdalommoduláció.
Ismeretes, hogy a morfin mikroinjekciója a középagy periaqueductalis szürkeállományába (PAG) (Tsou, Jang, 1964) (centrális szürkeállomány - CSV), valamint annak elektromos stimulációja (Reynolds, 1969) olyan mély fájdalomcsillapítást okoz, hogy még sebészeti beavatkozások sem okoznak észrevehető reakciókat. Amikor felfedezték az opioid receptorok és a természetes opiátok koncentrációs területeit, világossá vált, hogy az agytörzs ezen részei a supraspinalis leszálló modulációs vezérlőrendszerek közvetítőállomásai. Az egész rendszert, amint most világossá vált, a következőképpen mutatjuk be.
A B-endorfint transzmitterként használó sejtcsoport axonjai, amelyek a hipotalamusz nucl.arcuatus régiójában helyezkednek el (amely maga az agykéreg prefrontális és insuláris zónáinak irányítása alatt áll) keresztezi a periventrikuláris szürkeállományt az agykéregben. a harmadik kamra fala, amely a periaqueductalis szürkeállományban (PAG) végződik. Itt gátolják a lokális interneuronokat, így felszabadítják gátló hatásuk alól azokat a sejteket, amelyek axonjai lenyúlnak a nucleus raphe magnum régióba a medulla oblongata retikuláris képződésének közepén. Ennek a magnak a túlnyomóan szerotonerg (transzmitter - 5 - hidroxitriptamin) neuronjainak axonjai a gerincvelő dorsolaterális funiculusán lefelé irányulnak, és a hátszarv felszíni rétegeiben végződnek. A raphe spinalis axonok egy része és a retikuláris formációból származó axonok jelentős része noradrenerg. Így mind a szerotonerg, mind a noradrenerg neuronok az agytörzsben olyan struktúrákként működnek, amelyek blokkolják a nociceptív információt a gerincvelőben (Field, 1987). A biogén aminvegyületek jelenléte a fájdalomcsillapító rendszerekben magyarázza a triciklikus antidepresszánsok által okozott fájdalomcsillapítást. Ezek a gyógyszerek gátolják a szerotonin és a noradrenalin újrafelvételét a szinapszisban, és így fokozzák a transzmitterek gátló hatását a gerincvelő neuronjaira. A fájdalomérzékenység legerőteljesebb gátlását állatokban a nucl.raphe magnus (repcemag) közvetlen stimulálása okozza. Emberben a periventricularis és a periaqueductalis szürkeállomány a leggyakrabban használt beültethető elektródákkal végzett stimuláció a fájdalomcsillapítás céljából (Richardson, 1982). A fent említett spinothalamikus axonoktól a retikuláris formációig terjedő kollaterális magyarázhatja a heteroszegmentális akupunktúra hatását, mivel a gerinc nem specifikus neuronjait egy inger, például egy tűszúrás aktiválhatja (Bowsher, 1987).
Szomatikus és zsigeri érzékenység
Az érzékszervi érzések három fiziológiai osztályba sorolhatók: mechanoreceptív, hőfokÉs fájdalmas. A mechanoreceptív érzetek közé tartozik tapintható(érintés, nyomás, rezgés) és proprioceptív(testtartási) - testtartás, statikus helyzet és pozíció mozgás közben.
Az érzékenység az érzések keletkezésének helye szerint osztályozható exteroceptív(a test felszínéről származó érzések), zsigeri(a belső szervekben fellépő érzések) és mély(az érzések a mélyen fekvő szövetekből származnak - fascia, izmok, csontok).
· Szomatikus szenzoros jeleket nagy sebességgel, nagy pontossággal továbbítják a lokalizációt és meghatározzák az intenzitás minimális gradációit vagy az érzékelő jel erősségének változásait.
· Viscerális jeleket Alacsonyabb vezetési sebesség, a jelészlelés térbeli lokalizációjának kevésbé fejlett rendszere, az ingerlés erősségének kevésbé fejlett fokozatos rendszere és a jel gyors változásainak kevésbé közvetítő képessége jellemzi.
Szomatoszenzoros jeleket
Tapintható érzékenység
Az érintés, a nyomás és a rezgés tapintása az érzések külön típusa, de ugyanazok a receptorok érzékelik.
· Érzés érintés- a bőr és az alatta lévő szövetek érzékeny idegvégződéseinek stimulálásának eredménye.
· Érzés nyomás a mély szövetek deformációja következtében jelentkezik.
· Rezgés érzés gyors, ismétlődő érzékszervi ingerek eredményeként jelentkezik, amelyek ugyanazokra a receptorokra vonatkoznak, mint azok, amelyek érintést és nyomást érzékelnek.
Tapintási receptorok
Proprioceptívérzés
Az ebben a részben található anyagokat lásd a könyvben.
Átviteli útvonalak szomatoszenzoros jeleket
A testszegmensekből (ld. 9–8. ábra) szinte minden szenzoros információ a gerincvelői ganglionok szenzoros neuronjainak centrális folyamatain keresztül jut be a gerincvelőbe, áthaladva a hátgyökereken (9–2., 9–3. ábra). A gerincvelőbe jutva az érző neuronok központi folyamatai vagy közvetlenül a medulla oblongata-ba (lemniscalis rendszer: vékony vagy finom Gaulle fasciculus és cuneate fasciculus Burdach), vagy az interneuronokon végződnek, amelyek axonjai a thalamusba mennek. a ventrális, vagy elülső és laterális, vagy oldalsó spinothalamikus felszálló traktus része.
Rizs . 9-2. Gerincvelő . Kilátás hátulról. Magyarázatok a szövegben. A gerincvelő magjainak, lemezeinek és szakaszainak térképét lásd a 13. fejezet „A gerincvelő magjai és pályái” című részben.
· Vékony És ék alakú fürtök - vezetőképes módokon proprioceptív És tapintható érzékenység- a gerincvelő ugyanazon oldalának hátsó agyának részeként halad át, és a medulla oblongata vékony és sphenoid magjaiban végződik. Ezen magok neuronjainak axonjai a mediális hurok mentén (innen a név - lemniscal rendszer) az ellenkező oldalra mozognak, és a talamuszba mennek.
· Spinothalamikus pálya hasi- az ellenkező oldal elülső zsinórjában áthaladó projekciós afferens pálya. A gerinc ganglionokban elhelyezkedő első neuronok perifériás folyamatai végrehajtani tapintható És nyomógomb Érez tól től mechanoreceptorok bőr. Ezeknek a neuronoknak a központi folyamatai a hátgyökereken keresztül bejutnak a dorsalis funiculiokba, ahol 2-15 szegmensre emelkednek, és szinapszisokat képeznek a háti szarv interneuronjaival. Ezen neuronok axonjai az ellenkező oldalra mozognak, és továbbhaladnak az anterolaterális funicuszok elülső perifériás zónájában. Innen a pálya rostjai a thalamus posterolaterális ventrális magjába emelkednek a laterális spinothalamikus traktussal együtt.
· Spinothalamikus pálya oldalsó- az oldalsó zsinórban áthaladó projekciós afferens pálya. A perifériás receptorok a bőr szabad idegvégződései. A gerincvelői ganglionok pszeudounipoláris neuronjainak központi folyamatai a hátgyökerek oldalsó szakaszain keresztül bejutnak a gerincvelő ellentétes részébe, és a gerincvelőben 1-2 szegmenst felemelve szinapszisokat képeznek a neuronokkal. Rolandé kocsonyás anyagokat. Ezen neuronok axonjai valójában az oldalsó spinothalamikus traktust alkotják. Az ellenkező oldalra mennek, és az oldalsó zsinórok oldalsó szakaszaiban emelkednek. A spinothalamikus pályák áthaladnak az agytörzsön, és a thalamus ventrolateralis magjaiban végződnek. Ez fő- pálya véghezvitel fájdalmas És hőfok érzékenység.
Rizs . 9-3. Emelkedő ösvények érzékenység. A . A gerincvelői ganglionok szenzoros neuronjaitól (az első vagy elsődleges szenzoros neurontól) a második neuronokon (a gerincvelő interneuronjai vagy a sphenoid és a vékony velőmag idegsejtjei) át az út harmadik neuronjaiig - talamusz. Ezen neuronok axonjai az agykéregbe vetülnek. B . A különböző modalitást közvetítő neuronok elhelyezkedése a gerincvelő lemezeiben (római számok).
A hátsó zsinór vastag, myelinizált idegrostokból áll, amelyek 30-110 m/s sebességgel vezetik a jeleket; A spinothalamikus traktusok vékony, myelinizált rostokból állnak, amelyek néhány métertől 40 m/s-ig terjedő sebességgel vezetik az AP-kat.
Szomatoszenzoros ugat
Az ebben a részben található anyagokat lásd a könyvben.
Jelfeldolgozás növekvő vetületi utakon
Az ebben a részben található anyagokat lásd a könyvben.
Fájdalmas érzékenység
A fájdalom egy kellemetlen szenzoros és érzelmi érzés, amely tényleges vagy potenciális szövetkárosodáshoz kapcsolódik, vagy ilyen károsodással jellemezhető. A fájdalom egy védő jelzőmechanizmus a szervezet számára, és minden olyan szövetben előfordulhat, ahol a károsodás jelei megjelentek. A fájdalom gyors és lassú, akut és krónikus fájdalomra osztható.
· Gyors fájdalom 0,1 másodperccel a fájdalmas inger alkalmazása után érezhető. A gyors fájdalmat számos néven írják le: vágó, szúró, éles, elektromos stb. A fájdalomreceptoroktól a gerincvelőig a fájdalomjelek kis átmérőjű A rostok mentén továbbítódnak. d 6-30 m/s sebességgel.
· Lassú fájdalom 1 másodperc alatt vagy tovább lép fel, majd sok másodperc vagy perc alatt lassan növekszik (például lassú égő, tompa, lüktető, repeső, krónikus fájdalom). A lassú krónikus fájdalom a C rostok mentén 0,5-2 m/s sebességgel terjed.
A fájdalomjelek továbbítására szolgáló kettős rendszer megléte ahhoz a tényhez vezet, hogy az erős éles irritáció gyakran kettős fájdalomérzetet okoz. A gyors fájdalom azonnal átadódik, és egy második vagy egy kicsit később lassú fájdalom.
A fájdalom fogadása
A fájdalmat számos tényező okozza: mechanikai, termikus és kémiai fájdalomingerek. A gyors fájdalmat főként mechanikai és hőmérsékleti ingerek, lassú fájdalmat minden típusú inger generál. Egyes anyagok kémiai fájdalomcsillapítókként ismertek: káliumionok, tejsav, proteolitikus enzimek. A prosztaglandinok növelik a fájdalomvégződések érzékenységét, de közvetlenül nem gerjesztik őket. Fájdalomreceptorok ( nociceptorok) szabad idegvégződések (lásd 8-1A. ábra). Széles körben elterjedtek a bőr felületes rétegeiben, a csonthártyában, az ízületekben és az artériák falában. Más mély szövetekben kevesebb szabad idegvégződés van, de a kiterjedt szövetkárosodás a test szinte minden területén fájdalmat okozhat. A fájdalomreceptorok gyakorlatilag nem alkalmazkodnak.
· Akció kémiai ösztönzők, amely fájdalmat okoz, akkor nyilvánul meg, amikor a sérült szövetből kivonatot fecskendeznek be a bőr normál területére. A kivonat tartalmazza az összes fent leírt, fájdalmat okozó kémiai tényezőt. A legsúlyosabb fájdalmat az okozza , ami lehetővé tette, hogy a szövetkárosodás esetén a fájdalom fő okának tekintsék. Ezenkívül a fájdalom intenzitása korrelál a káliumionok helyi növekedésével és a proteolitikus enzimek aktivitásának növekedésével. A fájdalom megjelenése ebben az esetben a proteolitikus enzimek idegvégződésekre gyakorolt közvetlen hatásával és a membrán permeabilitásának növekedésével magyarázható a K számára. + , ami a fájdalom közvetlen oka.
· Szövet ischaemia, amely akkor jelentkezik, amikor a szövetben leáll a vérkeringés, néhány perc múlva erős fájdalmat okoz. Megfigyelték, hogy minél magasabb az anyagcsere a szövetben, annál gyorsabban jelentkezik a fájdalom, ha a véráramlás megszakad. Például egy mandzsetta felhelyezése a felső végtagon és a levegő pumpálása addig, amíg a véráramlás teljesen leáll, 15-20 másodperc múlva fájdalom jelentkezik a dolgozó izomban. Ugyanezen körülmények között a nem működő izom fájdalma néhány perccel később jelentkezik.
· Tejtermék sav. Az ischaemia során fellépő fájdalom lehetséges oka a nagy mennyiségű tejsav felhalmozódása, de nem kevésbé valószínű, hogy más kémiai tényezők (például proteolitikus enzimek) is képződnek a szövetben, és ez utóbbiak stimulálják a fájdalom idegvégződéseit. .
· Izmos görcs fájdalomhoz vezet, amely számos klinikai fájdalomszindróma hátterében áll. A fájdalom oka lehet a görcs közvetlen hatása az izmok mechanoszenzitív fájdalomreceptoraira. Valószínűbb, hogy a fájdalom oka az izomgörcs közvetett hatása, amely összenyomja az ereket és ischaemiát okoz. Végül, a görcs növeli az anyagcsere-folyamatok sebességét az izomszövetben, feltételeket teremtve az ischaemia hatásának fokozásához és a fájdalmat okozó anyagok felszabadulásához.
· Fájdalmas receptorok gyakorlatilag Nem alkalmazkodni. Egyes esetekben a fájdalomreceptorok gerjesztése nemcsak nem csökken, hanem folyamatosan növekszik (például tompa ívelő fájdalom formájában). A fájdalomreceptorok fokozott érzékenységét ún hiperalgézia. A fájdalomérzékenység küszöbének csökkenése észlelhető hosszan tartó hőmérséklet-stimulációval. A nociceptorok alkalmazkodóképességének hiánya nem teszi lehetővé az alanynak, hogy megfeledkezzen a fájdalmas ingerek káros hatásairól teste szöveteire.
Fájdalomjelek továbbítása
A gyors és lassú fájdalom saját idegpályájuknak felel meg: pálya véghezvitel gyors fájdalom És pálya véghezvitel lassú krónikus fájdalom.
Gyors fájdalom végrehajtása
A gyors fájdalom (9-7A. ábra) receptorokról történő levezetését Ad típusú rostok végzik, amelyek a háti gyökerek mentén belépnek a gerincvelőbe, és szinaptikusan érintkeznek az ugyanazon oldali hátszarv idegsejtjeivel. Az ugyanazon az oldalon lévő másodrendű neuronokkal szinapszisok kialakulása után az idegrostok az ellenkező oldalra mozdulnak el, és az anterolateralis zsinórokban a spinothalamikus traktus részeként felemelkednek az agytörzsig. Az agytörzsben egyes rostok szinaptikusan érintkeznek a retikuláris formáció neuronjaival, míg a rostok zöme a thalamusba jut, a ventro-bazális komplexben végződve a lemniscalis rendszer rostjaival együtt, amelyek érintési érzékenységet hordoznak. A rostok egy kis része a thalamus hátsó magjaiban végződik. Ezekről a thalamicus területekről a jelek más bazális agyi struktúrákhoz és a szomatoszenzoros kéreghez jutnak (9-7A ábra).
Rizs . 9-7. A fájdalom átvitelének útjai érzékenység(A) és antinociceptív rendszer (B).
· Lokalizáció gyors fájdalom a test különböző részein jobban megkülönböztethető, mint a lassú krónikus fájdalom.
· Adás fájdalmas impulzusok(9–7B, 9–8. ábra). A glutamát a fájdalomingerek átvitelében serkentő neurotranszmitterként vesz részt a gerincvelői ganglion szenzoros neuronjainak központi folyamatai és a spinothalamikus traktus neuronjainak perikarionjai közötti szinapszisokban. A P szubsztancia szekréciójának blokkolása és a fájdalom csillapítása a szenzoros neuron központi folyamata terminálisának membránjába épített opioid peptid receptorokon keresztül valósul meg (példa a preszinaptikus gátlás jelenségére). Az opioid peptid forrása az interneuron.
Rizs . 9–8. A fájdalomimpulzusok útja (nyilak). A P szubsztancia a gerjesztést a szenzoros neuron központi folyamatából a spinothalamikus traktus idegsejtjébe továbbítja. Az opioid receptorokon keresztül az interneuronból származó enkefalin gátolja a P anyag szenzoros neuronból történő kiválasztását és a fájdalomjelek átvitelét.[ 11 ].
Lassú krónikus fájdalom kezelése
A szenzoros neuronok központi folyamatai a laminae II és III neuronjain végződnek. A második neuronok hosszú axonjai átjutnak a gerincvelő másik oldalára, és az anterolateralis agyvelő részeként felszállnak az agyba. Ezek a rostok, amelyek a paleospinothalamikus traktus részeként a lassú krónikus fájdalom jeleit hordozzák, kiterjedt szinaptikus kapcsolatokkal rendelkeznek az agytörzsben, a medulla oblongata retikuláris magjaiban, a hídon és a középagyban, a thalamusban, a tegmentalis területen és az agyban végződnek. a Sylvius vízvezetékét körülvevő szürkeállomány. Az agytörzsből fájdalomjelek érkeznek a thalamus intraplate és ventrolateralis magjaiba, a hipotalamuszba és más agyalapi struktúrákba (9-7B. ábra).
· Lokalizáció lassú krónikus fájdalom. A lassú krónikus fájdalom nem a test egyes pontjain lokalizálódik, hanem annak nagy részein, mint a kar, láb, hát stb. Ezt a lassú fájdalmat vezető utak poliszinaptikus, diffúz kapcsolatai magyarázzák.
· Központi fokozat lassú fájdalom. Az állatok szomatoszenzoros kéregének teljes eltávolítása nem rontja a fájdalomérzékelési képességüket. Ezért az agytörzs retikuláris képződményén, a talamuszon és más mögöttes központokon keresztül az agyba jutó fájdalomimpulzusok a fájdalom tudatos érzékelését idézhetik elő. A szomatoszenzoros kéreg részt vesz a fájdalom minőségének felmérésében.
· Neurotranszmitter lassú fájdalom a C-szálak végein - . A gerincvelőbe belépő C típusú fájdalomrostok néhány ezredmásodperc alatt felszabadítják a neurotranszmittereket, a glutamátot és a P anyagot. A P anyag lassabban szabadul fel, és másodperceken vagy akár perceken belül eléri hatásos koncentrációját.
Fájdalomcsillapító rendszer
Az emberi szervezet nemcsak érzékeli és meghatározza a fájdalomjelek erősségét és minőségét, hanem képes csökkenteni, sőt elnyomni a fájdalomrendszerek aktivitását. A fájdalomra adott egyéni válaszok köre szokatlanul széles, és a fájdalomra adott válasz nagymértékben függ attól, hogy az agy az antinociceptív (fájdalomcsillapító, fájdalomcsillapító) rendszer segítségével elnyomja-e az idegrendszerbe jutó fájdalomjeleket. Az antinociceptív rendszer (9-7B. ábra) három fő összetevőből áll.
1 . Összetett fékezés fájdalom, a gerincvelő hátsó szarvaiban található. Itt a fájdalom blokkolva van, mielőtt elérné az agy befogadó részeit.
2 . Nagy mag varrás, a híd és a medulla oblongata közötti középvonalban helyezkedik el; retikuláris paragiáns sejt mag, amely a medulla oblongata oldalsó részében található. Ezekből a magokból a jelek a posterolaterális oszlopok mentén haladnak a gerincvelőbe.
3 . Okolovoprovodnoe szürke anyag És periventricularis vidék a középagy és a felső híd, amely körülveszi Sylvius vízvezetékét és a harmadik és negyedik kamra egyes részeit. Az ezekről a fájdalomcsillapító területekről származó neuronok jeleket küldenek a raphe nucleus magnusnak és a reticularis paragiant sejtmagnak.
A periaqueductalis szürkeállomány vagy a raphe nuclei magnus elektromos stimulációja szinte teljesen elnyomja a gerincvelő hátsó gyökerein áthaladó fájdalomjeleket. A fedő agyi struktúrák stimulálása viszont gerjeszti a periventricularis magokat és a hypothalamus előagyi medialis fasciculumát, és ezáltal fájdalomcsillapító hatást vált ki.
· Neurotranszmitterek antinociceptív rendszerek. A fájdalomcsillapító rendszer idegrostjainak végződésein felszabaduló mediátorok és. A fájdalomcsillapító rendszer különböző részei érzékenyek a morfiumra, az opiátokra és az opioidokra ( b -endorfin, enkefalin, dinorfin). Különösen enkefalinokat és dinorfint találtak az agytörzs és a gerincvelő fájdalomcsillapító rendszerének szerkezetében.
A szinapszisokat tartalmazó idegrostok szinapszisokat képeznek a raphe fő mag neuronjaival. Ezen neuronok axonjai a gerincvelő hátsó szarvában végződnek, és a végződéseikből válnak ki. A szerotonin pedig gerjeszti az enkefalinerg neuronokat a gerincvelő hátsó szarvában (9-8. ábra). Az enkefalin preszinaptikus gátlást és posztszinaptikus gátlást okoz a C és A típusú fájdalomrost szinapszisokban d a gerincvelő hátsó szarvaiban. Feltételezhető, hogy a preszinaptikus gátlás az idegvégződések membránjában lévő kalciumcsatornák blokkolásának eredményeként következik be.
Központi fékezés És zavaró irritáció
· A fájdalomcsillapító rendszer aktiválása szempontjából az a jól ismert tény, hogy a sebesültek harc közben elfelejtik a fájdalmat (stressz fájdalomcsillapítás), valamint a fájdalom csökkentése a sérült testterület simogatása vagy rezgése során, sokak számára ismert tény. személyes tapasztalat, magyarázzák.
· A fájdalmas terület elektromos vibrátorral történő stimulálása némi fájdalomcsillapítást is nyújt. Az akupunktúrát több mint 4000 éve használják a fájdalom megelőzésére vagy enyhítésére, és bizonyos esetekben az akupunktúrát nagy sebészeti beavatkozások elvégzésére használják.
· A fájdalomjelek gátlása a központi szenzoros pályákban szintén megmagyarázhatja a belső szervi gyulladásos területen a bőr stimulálására használt zavaró stimuláció hatékonyságát. Tehát a mustárvakolat és a paprikatapasz ezen az elven működik.
Utalt fájdalom
A belső szervek irritációja gyakran okoz fájdalmat, amely nemcsak a belső szervekben, hanem egyes szomatikus struktúrákban is érezhető, amelyek a fájdalom okának helyétől meglehetősen távol helyezkednek el. Ezt a fajta fájdalmat utalásnak (sugárzónak) nevezik.
A hivatkozott fájdalom legismertebb példája a szívfájdalom, amely a bal karba sugárzik. A leendő orvosnak azonban tudnia kell, hogy a fájdalom tükröződési területei nem sztereotipek, és a reflexió szokatlan területei gyakran megfigyelhetők. A szívfájdalom például lehet pusztán hasi, kisugározhat a jobb karba, sőt a nyakba is.
Szabály dermatomerek . A bőr, az izmok, az ízületek és a belső szervek afferens rostjai a háti gyökerek mentén, meghatározott térbeli rendben bejutnak a gerincvelőbe. Az egyes hátgyökerekből származó bőr afferens rostok egy korlátozott bőrterületet beidegznek, amelyet dermatomernek neveznek (9-9. ábra). A hivatkozott fájdalom általában ugyanazon embrionális szegmensből vagy dermatomerből fejlődő struktúrákban jelentkezik. Ezt az elvet „dermatomer szabálynak” nevezik. Például a szív és a bal kar azonos szegmentális jellegű, és a here idegellátásával az urogenitális gerincről vándorolt, ahonnan a vesék és az ureterek keletkeztek. Ezért nem meglepő, hogy az ureterből vagy a veséből származó fájdalom a herékbe sugárzik.
Rizs . 9-9. Bőrgyógyászok
Konvergencia és enyhülés a hivatkozott fájdalom mechanizmusában
A hivatkozott fájdalom kialakulásában nemcsak az idegrendszerbe egy-egy szegmentális szinten bekerülő zsigeri és szomatikus idegek, hanem a spinothalamikus traktusokon áthaladó nagyszámú érzőidegrost is részt vesz. Ez megteremti a feltételeket a perifériás afferens rostok konvergenciájához a talamusz neuronokon, azaz. szomatikus és zsigeri afferensek ugyanazon neuronokon konvergálnak (9-10. ábra).
· Elmélet konvergencia. A szomatikus fájdalomra vonatkozó információk nagyobb sebessége, konzisztenciája és gyakorisága segíti az agyat abban, hogy megszilárdítsa azokat az információkat, amelyek szerint a megfelelő idegpályákba belépő jeleket a test bizonyos szomatikus területein fájdalmas ingerek okozzák. Ha ugyanazokat az idegpályákat gerjeszti a zsigeri fájdalom afferens rostjainak aktivitása, az agyba érkező jel nem differenciálódik, és a fájdalom a test szomatikus területére vetül.
· Elmélet megkönnyebbülés. Az utalt fájdalom eredetének egy másik elmélete (az úgynevezett megkönnyebbülés elmélete) azon a feltételezésen alapul, hogy a belső szervekből érkező impulzusok csökkentik a spinothalamikus neuronok küszöbét a szomatikus területekről érkező afferens fájdalomjelek hatására.. Megkönnyebbülés esetén a szomatikus terület minimális fájdalomaktivitása is átjut az agyba.
Rizs . 9-10. Utalt fájdalom
Ha a konvergencia az egyetlen magyarázat a hivatkozott fájdalom eredetére, akkor a hivatkozott fájdalom területének helyi érzéstelenítése nem befolyásolhatja a fájdalmat. Másrészt, ha küszöb alatti csillapító hatások is szerepet játszanak az utalt fájdalom előfordulásában, akkor a fájdalomnak el kell tűnnie. A helyi érzéstelenítés hatása a hivatkozott fájdalom területére változó. Az erős fájdalom általában nem múlik el, a mérsékelt fájdalom teljesen megszűnhet. Ezért mindkét tényező az konvergencia És megkönnyebbülés- részt venni az utalt fájdalom előfordulásában.
Szokatlan és hosszan tartó fájdalom
Néhány embernél a perifériás idegeket érintő károsodások és betegségek súlyos, legyengítő és rendellenesen tartós fájdalmat okoznak.
· Hiperalgézia, amelyben az általában mérsékelt fájdalomérzethez vezető ingerek súlyos, hosszan tartó fájdalmat okoznak.
· Kauzalgia- tartós égő érzés, amely általában a perifériás ideg érzőrostjainak érkárosodása után alakul ki.
· Allodynia- fájdalmas érzések, amelyekben a semleges ingerek (például enyhe lélegzetvétel vagy a ruházat érintése) heves fájdalmat okoznak.
· Hiperpátia- olyan fájdalmas érzés, amelyben a fájdalomküszöb megemelkedik, de annak elérésekor intenzív, égető fájdalom lép fel.
· Fantom a fájdalom egy hiányzó végtag fájdalmas érzése.
Ezeknek a fájdalomszindrómáknak az okai nem teljesen tisztázottak, de ismert, hogy az ilyen típusú fájdalmak nem csillapíthatók helyi érzéstelenítéssel vagy idegvágással. Kísérleti vizsgálatok azt mutatják, hogy az idegkárosodás a noradrenerg idegrostok intenzív proliferációjához és elágazásához vezet a szenzoros ganglionokban, ahonnan a háti gyökerek a sérült terület felé emelkednek. Nyilvánvalóan a szimpatikus kisülések hozzájárulnak a szokatlan fájdalomjelek megjelenéséhez. Így egy ördögi kör alakul ki a periférián. A vele kapcsolatos sérült idegrostokat noradrenalin stimulálja a háti gyökerek szintjén. a -Adrenerg blokád csökkenti a fájdalmas ok-okozati érzéseket.
Thalamic szindróma. Spontán fájdalom jelentkezhet a thalamus szintjén. Thalamus-szindrómában a hátulsó talamuszmagok károsodása következik be, amelyet általában a hátsó agyi artéria ágainak elzáródása okoz. Az ebben a szindrómában szenvedő betegek hosszan tartó, súlyos, rendkívül kellemetlen fájdalom rohamokat tapasztalnak, amelyek spontán módon vagy különféle szenzoros ingerekre reagálva jelentkeznek.
A fájdalom megfelelő dózisú fájdalomcsillapítók alkalmazásával csillapítható, de ez nem minden esetben történik meg. Az elviselhetetlen fájdalom enyhítésére a hátsó gyökerek krónikus irritációját alkalmazzák beültetett elektródákkal. Az elektródák egy hordozható stimulátorhoz csatlakoznak, a páciens szükség esetén saját magát stimulálja. A fájdalom enyhülését nyilvánvalóan az impulzusok antidromikus vezetése éri el a kollaterálisokon keresztül a hátsó gyökerek fájdalomcsillapító rendszeréhez. A periaqueductalis szürkeállomány önstimulációja szintén segít csökkenteni az elviselhetetlen fájdalmat, valószínűleg a felszabadulás miatt.
Visceralis fájdalom
A gyakorlati gyógyászatban a belső szervekben fellépő fájdalom a gyulladások, fertőző betegségek és egyéb rendellenességek fontos tünete. Minden olyan inger, amely túlzottan stimulálja a belső szervek idegvégződéseit, fájdalmat okoz. Ezek közé tartozik a zsigeri szövet ischaemia, a belső szervek felületének kémiai károsodása, az üreges szervek simaizmainak görcse, az üreges szervek megnyúlása és a szalagos apparátus megnyúlása. A zsigeri fájdalom minden fajtája az autonóm idegeken áthaladó, főleg szimpatikus idegrostokon keresztül terjed. A fájdalomrostokat vékony C-rostok képviselik, amelyek krónikus fájdalmat vezetnek.
A zsigeri fájdalom okai
· Ischaemia fájdalmat okoz a savas anyagcseretermékek és szöveti bomlástermékek, valamint a fájdalom idegvégződéseit irritáló proteolitikus enzimek képződése következtében.
· Görcs üreges szervek(például bélszakasz, húgycső, epehólyag, epeutak stb.) a fájdalomreceptorok mechanikai irritációját okozza. Néha a mechanikai irritációt görcs okozta ischaemiával kombinálják. A görcsös szervből származó fájdalom gyakran akut görcsös roham formájában jelentkezik, bizonyos mértékig fokozódik, majd fokozatosan csökken.
· Kémiai irritáció olyan esetekben fordulhat elő, amikor a gyomor-bél traktusból káros anyagok kerülnek a hasüregbe. A gyomornedv bejutása a hasüregbe kiterjed a fájdalomreceptorok irritációjának széles területére, és elviselhetetlenül akut fájdalmat okoz.
· Túlnyújtás üreges szervek mechanikusan irritálja a fájdalomreceptorokat, és megzavarja a véráramlást a szerv falában.
Fejfájás
A fejfájás egyfajta utalt fájdalom, amelyet fájdalmas érzésként érzékelnek, amely a fej felszínén jelentkezik. A fájdalom sok fajtája a koponyán belüli fájdalmas ingerekből, mások a koponyán kívüli ingerekből erednek.
Fejfájás koponyán belüli eredet
· Érzékeny Nak nek fájdalom vidék belül koponyák. Maga az agy teljesen mentes a fájdalomérzékenységtől. Még a szenzoros kéreg bemetszése vagy elektromos stimulációja is csak véletlenül okozhat fájdalmat. A szomatoszenzoros kéregben képviselt területeken jelentkező fájdalom helyett enyhe bizsergő érzés jelentkezik - paresztézia. Ezért nem valószínű, hogy a legtöbb fejfájást az agy parenchyma károsodása okozza.
· Nyomás tovább vénás melléküregek Az agyat körülvevő tentorium sérülése vagy a dura mater megnyúlása az agy alján heves fájdalmat okozhat, amelyet fejfájásként határoznak meg. Minden típusú trauma (az agyhártya ereinek zúzódása, nyújtása, csavarása) fejfájást okoz. Különösen érzékenyek a középső agyi artéria szerkezetei.
· Meningeális fájdalom- a fejfájás legsúlyosabb típusa, amely az agyhártya gyulladásos folyamatai során jelentkezik, és a fej teljes felületén tükröződik.
· Fájdalom nál nél csökken nyomás az agy-gerincvelői folyadékban a folyadék mennyiségének csökkenése és az agyhártya megnyúlása miatt az agy súlya miatt következik be.
· Fájdalom nál nél migrén spasztikus vaszkuláris reakciók eredményeként jelentkezik. Úgy gondolják, hogy a migrén hosszan tartó érzelmek vagy stressz következtében alakul ki, amelyek a fej bizonyos artériás ereiben görcsöt okoznak, beleértve az agyat ellátókat is. A görcs okozta ischaemia következtében az érfal tónusának elvesztése következik be, amely 24-48 óráig tart. A vérnyomás impulzusingadozása intenzívebben feszíti meg az artériák elernyedt atonikus érfalait, és az artériák falainak túlfeszítése, beleértve az extracranialisakat is (például a temporális artériákat), fejfájáshoz vezet.
A migrén eredetét érzelmi rendellenességek is magyarázzák, amelyek a kérgi depresszió terjedéséhez vezetnek. A depresszió a kálium-ionok lokális felhalmozódását okozza az agyszövetben, ami vaszkuláris görcsöt okoz.
· Alkohol függő fájdalom az acetaldehid agyhártyára gyakorolt közvetlen mérgező irritáló hatása okozza.
Extracranialis eredetű fejfájás
· Fejeket fájdalom V eredmény izmos görcs akkor fordul elő, ha érzelmi feszültség van a koponyához és a vállövhöz kapcsolódó számos izomban. A fájdalom a fej felszínén tükröződik, és koponyán belüli fájdalomhoz hasonlít.
· Fejeket fájdalom nál nél irritáció orr üregek És alárendelt mellékmondatok melléküregek orr nem nagy intenzitásúak, és a fej elülső felületén tükröződnek.
· Fejeket fájdalom nál nél jogsértések funkciókat szem a ciliáris izom erős összehúzódásainál fordulhat elő, amikor jobb látást próbálnak elérni. Ez az arc és a külső szemizmok reflexgörcsét és fejfájást okozhat. A második típusú fájdalom akkor figyelhető meg, amikor a retinát ultraibolya sugárzás „égeti”, valamint a kötőhártya irritációját.



