A vér összetétele és funkciói. Immunitás. A vér összetétele és az immunrendszer Az immunrendszer funkciói
>>anatómia és élettan
Immunitás(latin immunitas - valamitől megszabadulni) olyan élettani funkció, amely immunissá teszi a szervezetet az idegen antigénekkel szemben. Az emberi immunitás immunissá teszi számos baktérium, vírus, gomba, féreg, protozoa és különféle állati méreg ellen. Ezenkívül az immunrendszer megvédi a szervezetet a rákos sejtektől.
Az immunrendszer feladata minden idegen struktúra felismerése és elpusztítása. Idegen szerkezettel érintkezve az immunrendszer sejtjei immunválaszt váltanak ki, ami az idegen antigén eltávolításához vezet a szervezetből.
Az immunitás működését a szervezet immunrendszerének munkája biztosítja, amely különböző típusú szerveket és sejteket foglal magában. Az alábbiakban részletesebben megvizsgáljuk az immunrendszer felépítését és működésének alapelveit.
Az immunrendszer anatómiája
Az immunrendszer anatómiája rendkívül heterogén. Általában az immunrendszer sejtjei és humorális tényezői a test szinte minden szervében és szövetében jelen vannak. Kivételt képeznek a szem egyes részei, férfiaknál a herék, a pajzsmirigy, az agy – ezeket a szerveket szöveti gát védi az immunrendszertől, ami szükséges a normál működésükhöz.
Általánosságban elmondható, hogy az immunrendszer működését kétféle tényező biztosítja: sejtes és humorális (azaz folyékony). Az immunrendszer sejtjei (különböző típusú leukociták) keringenek a vérben és bejutnak a szövetekbe, folyamatosan figyelemmel kísérve a szövetek antigén összetételét. Emellett nagy számban keringenek a vérben különböző antitestek (humorális, fluid faktorok), amelyek szintén képesek idegen struktúrák felismerésére és elpusztítására.
Az immunrendszer felépítésében megkülönböztetünk központi és perifériás struktúrákat. Az immunrendszer központi szervei a csontvelő és a csecsemőmirigy (csecsemőmirigy). A csontvelőben (vörös csontvelő) az immunrendszer sejtjeinek kialakulása az ún őssejtek, amelyekből minden vérsejt (vörösvértestek, leukociták, vérlemezkék) keletkezik. A csecsemőmirigy (csecsemőmirigy) a mellkasban, közvetlenül a szegycsont mögött található. A csecsemőmirigy gyermekeknél jól fejlett, de az életkor előrehaladtával involúción megy keresztül, és felnőtteknél gyakorlatilag hiányzik. A csecsemőmirigyben a limfociták – az immunrendszer specifikus sejtjei – differenciálódása megy végbe. A differenciálódás folyamatában a limfociták „megtanulják” felismerni „saját” és „idegen” struktúrájukat.
Az immunrendszer perifériás szervei nyirokcsomók, lép és nyirokszövet képviseli (ilyen szövet található például a palatinus mandulákban, a nyelv gyökerében, a nasopharynx hátsó falán, a belekben).
A nyirokcsomók Ezek limfoid szövetek gyűjteményei (valójában az immunrendszer sejtjeinek gyűjteménye), amelyeket egy membrán vesz körül. A nyirokcsomók nyirokereket tartalmaznak, amelyeken keresztül a nyirok áramlik. A nyirokcsomó belsejében a nyirok megszűrik és megtisztítják az összes idegen struktúrát (vírusok, baktériumok, rákos sejtek). A nyirokcsomót elhagyó erek egy közös csatornába egyesülnek, amely egy vénába áramlik.
Lép nem más, mint egy nagy nyirokcsomó. Felnőtt emberben a lép tömege a szervben felhalmozódott vér mennyiségétől függően akár több száz grammot is elérhet. A lép a hasüregben, a gyomortól balra található. Naponta nagy mennyiségű vér pumpálódik át a lépen, amely a nyirokcsomók nyirokrendszeréhez hasonlóan szűrés és tisztításon esik át. Ezenkívül bizonyos mennyiségű vér tárolódik a lépben, amelyre a szervezetnek jelenleg nincs szüksége. Fizikai aktivitás vagy stressz során a lép összehúzódik, és vért bocsát ki az erekbe, hogy kielégítse a szervezet oxigénszükségletét.
Nyirokszövet szétszórva a testben kis csomók formájában. A limfoid szövet fő funkciója a helyi immunitás biztosítása, ezért a limfoid szövetek legnagyobb felhalmozódása a szájban, a garatban és a belekben található (a test ezen területeit gazdagon népesítik be különféle baktériumok).
Ezen kívül a különböző szervekben ún mesenchymalis sejtek, amely immunfunkciót tud ellátni. Sok ilyen sejt található a bőrben, a májban és a vesében.
Az immunrendszer sejtjei
Az immunrendszer sejtjeinek általános neve leukociták. A leukocita család azonban nagyon heterogén. A leukociták két fő típusát különböztetjük meg: szemcsés és nem szemcsés.
Neutrophilek- a leukociták legtöbb képviselője. Ezek a sejtek egy megnyúlt sejtmagot tartalmaznak, amely több szegmensre oszlik, ezért néha szegmentált leukocitáknak nevezik őket. Mint az immunrendszer minden sejtje, a neutrofilek a vörös csontvelőben képződnek, és érés után a vérbe kerülnek. A neutrofilek keringési ideje a vérben nem hosszú. Néhány órán belül ezek a sejtek áthatolnak az erek falán, és bejutnak a szövetbe. A szövetekben eltöltött idő után a neutrofilek visszatérhetnek a vérbe. A neutrofilek rendkívül érzékenyek a gyulladás jelenlétére a szervezetben, és képesek a gyulladt szövetekbe irányítottan vándorolni. A szövetbe jutva a neutrofilek megváltoztatják alakjukat - kerekből elágazóvá válnak. A neutrofilek fő funkciója a különböző baktériumok semlegesítése. A szöveteken való mozgáshoz a neutrofil sajátos lábakkal van felszerelve, amelyek a sejt citoplazmájának kinövései. A baktérium felé haladva a neutrofil körülveszi folyamataival, majd speciális enzimek segítségével „lenyeli” és megemészti. Az elhalt neutrofilek a gyulladásos területeken (például a sebekben) genny formájában halmozódnak fel. A vérben lévő neutrofilek száma megnövekszik a különféle bakteriális jellegű gyulladásos betegségek során.
Basophilok aktívan részt venni az azonnali allergiás reakciók kialakulásában. A szövetbe jutva a bazofilek hízósejtekké alakulnak, amelyek nagy mennyiségű hisztamint tartalmaznak, egy biológiailag aktív anyagot, amely serkenti az allergia kialakulását. A bazofileknek köszönhetően a rovarok vagy állatok mérgei azonnal elzáródnak a szövetekben, és nem terjednek el az egész testben. A bazofilek a heparin segítségével szabályozzák a véralvadást is.
Limfociták. A limfocitáknak többféle típusa létezik: B-limfociták (B-limfociták), T-limfociták (T-limfociták), K-limfociták (K-limfociták), NK-limfociták (természetes gyilkos sejtek). ) és a monociták .
B limfociták felismerik az idegen struktúrákat (antigéneket), miközben specifikus antitesteket (idegen struktúrák ellen irányuló fehérjemolekulákat) termelnek.
T limfociták ellátja az immunitás szabályozásának funkcióját. A T-helperek serkentik az antitestek termelését, a T-szuppresszorok pedig gátolják azt.
K limfociták képes elpusztítani az antitestekkel jelölt idegen struktúrákat. Ezeknek a sejteknek a hatására különböző baktériumok, rákos sejtek vagy vírusokkal fertőzött sejtek pusztulhatnak el.
NK limfociták ellenőrizni kell a testsejtek minőségét. Ugyanakkor az NK limfociták képesek elpusztítani azokat a sejteket, amelyek tulajdonságaik különböznek a normál sejtektől, például a rákos sejtektől.
Monociták Ezek a legnagyobb vérsejtek. A szövetbe jutva makrofágokká alakulnak. A makrofágok nagy sejtek, amelyek aktívan elpusztítják a baktériumokat. A makrofágok nagy mennyiségben halmozódnak fel a gyulladásos területeken.
A neutrofilekhez képest (lásd fent) a limfociták bizonyos típusai aktívabbak a vírusokkal szemben, mint a baktériumok, és nem pusztulnak el az idegen antigénnel való reakció során, ezért nem képződik genny a vírusok által okozott gyulladásos területeken. A limfociták a krónikus gyulladásos területeken is felhalmozódnak.
A leukocita populáció folyamatosan megújul. Minden másodpercben milliónyi új immunsejt képződik. Egyes immunrendszer sejtjei csak néhány óráig élnek, míg mások több évig is fennmaradhatnak. Ez az immunitás lényege: ha egyszer találkozik egy antigénnel (vírussal vagy baktériummal), az immunsejt „emlékezik” rá, és a következő találkozáskor gyorsabban reagál, a szervezetbe jutás után azonnal blokkolja a fertőzést.
Egy felnőtt emberi test immunrendszerének szerveinek és sejtjeinek össztömege körülbelül 1 kilogramm. Az immunrendszer sejtjei közötti kölcsönhatások rendkívül összetettek. Általánosságban elmondható, hogy az immunrendszer különböző sejtjeinek összehangolt munkája biztosítja a szervezet megbízható védelmét a különféle fertőző ágensekkel és saját mutáns sejtjeivel szemben.
Az immunsejtek védő funkciójukon túl szabályozzák a testsejtek növekedését és szaporodását, valamint a gyulladásos területeken a szövetek helyreállítását.
Az emberi szervezetben az immunrendszer sejtjein kívül számos nem specifikus védekező faktor is létezik, amelyek az úgynevezett fajimmunitást alkotják. Ezeket a védőfaktorokat a komplementrendszer, a lizozim, a transzferrin, a C-reaktív fehérje, az interferonok képviselik.
Lizozim egy speciális enzim, amely elpusztítja a baktériumok falát. A lizozim nagy mennyiségben található a nyálban, ami megmagyarázza antibakteriális tulajdonságait.
Transzferin egy olyan fehérje, amely a baktériumokkal versenyez bizonyos, a fejlődésükhöz szükséges anyagok (például vas) felfogásában. Ennek eredményeként a baktériumok növekedése és szaporodása lelassul.
C-reaktív protein bókként aktiválódik, amikor idegen struktúrák kerülnek a vérbe. Ennek a fehérjének a baktériumokhoz való kötődése sebezhetővé teszi őket az immunrendszer sejtjeivel szemben.
Interferonok- Ezek összetett molekuláris anyagok, amelyeket a sejtek a vírusok szervezetbe való behatolására válaszul bocsátanak ki. Az interferonoknak köszönhetően a sejtek immunissá válnak a vírussal szemben.
Bibliográfia:
- Khaitov R.M. Immunogenetika és immunológia, Ibn Sina, 1991
- Leszkov, V.P. Klinikai immunológia orvosoknak, M., 1997
- Boriszov L.B. Orvosi Mikrobiológia, Virológia, Immunológia, M.: Medicina, 1994
Az oldal csak tájékoztató jellegű hivatkozási információkat tartalmaz. A betegségek diagnosztizálását és kezelését szakember felügyelete mellett kell elvégezni. Minden gyógyszernek van ellenjavallata. Szakorvosi konzultáció szükséges!
Írásban -POSITIV- Olvassa el az idézett üzenetet
Miből áll a vér és hogyan működik az immunrendszer?Az immunrendszer funkciói
Az immunrendszer fő feladata a szervezet makromolekuláris és sejtállandóságának felügyelete, megvédve a szervezetet minden idegentől. Az immunrendszer az idegrendszerrel és az endokrin rendszerrel együtt szabályozza és irányítja a szervezet összes fiziológiai reakcióját, ezáltal biztosítja a szervezet létfontosságú tevékenységét és életképességét. Az immunkompetens sejtek a gyulladásos reakció lényeges elemei, és nagymértékben meghatározzák annak természetét és lefolyását. Az immunkompetens sejtek fontos funkciója a szöveti regenerációs folyamatok szabályozása és szabályozása.
Az immunrendszer fő funkcióját specifikus (immun) reakciók kifejlesztésén keresztül látja el, amelyek alapja az „ön” és az „idegen” felismerésének képessége, majd az idegen eltávolítása. A humorális immunitás alapját az immunreakció eredményeként megjelenő specifikus antitestek képezik, a sejtes immunitás fő hordozói a szenzitizált limfociták.Az immunrendszerre jellemző az „immunológiai memória” jelensége, amelyre az jellemző, hogy az antigénnel való ismételt érintkezés az immunválasz felgyorsult és fokozott kialakulását idézi elő, ami az elsődleges immunválaszhoz képest hatékonyabb védelmet biztosít a szervezet számára. A másodlagos immunválasznak ez a sajátossága alapozza meg a vakcinázás indokát, amely sikeresen véd a legtöbb fertőzés ellen. Meg kell jegyezni, hogy az immunreakciók nem mindig csak védő szerepet töltenek be, hanem a szervezetben zajló immunpatológiai folyamatok okai lehetnek, és számos emberi szomatikus betegséget okozhatnak.
Az immunrendszer felépítése
Az emberi immunrendszert limfomieloid szervek és limfoid szövetek komplexe képviseli, amelyek a légzőrendszerhez, az emésztőrendszerhez és a húgyúti rendszerhez kapcsolódnak. Az immunrendszer szervei a következők: csontvelő, csecsemőmirigy, lép, nyirokcsomók. Az immunrendszerhez a felsorolt szerveken kívül a nasopharynx mandulája, a bél limfoid (Peyer-féle) foltjai, a gyomor-bél traktus nyálkahártyájában található számos nyirokcsomó, légzőcső, urogenitális traktus, diffúz limfoid szövet is tartozik. , valamint a bőr limfoid sejtjei és az interepiteliális limfociták.
Az immunrendszer fő elemei a limfoid sejtek. Az emberi limfociták teljes száma 1012 sejt. Az immunrendszer második fontos eleme a makrofágok. Ezeken a sejteken kívül a granulociták részt vesznek a szervezet védekező reakcióiban. A limfoid sejtek és a makrofágok az immunkompetens sejtek fogalma alatt egyesülnek.
Az immunrendszer T-linkre és B-linkre vagy T-immunrendszerre és B-immunrendszerre oszlik. A T-immunrendszer fő sejtjei a T-limfociták, a B-immunrendszer fő sejtjei a B-limfociták. Az immunrendszer T-rendszerének fő szerkezeti képződményei közé tartozik a csecsemőmirigy, a lép T-zónái és a nyirokcsomók; B-rendszerek immunitás - csontvelő, B-zónák a lép (reprodukciós központok) és a nyirokcsomók (kérgi zóna). Az immunrendszer T-linkje a sejtes típusú reakciókért, az immunrendszer B-linkje humorális típusú reakciókat valósít meg. A T-rendszer vezérli és szabályozza a B-rendszer működését. A B-rendszer viszont befolyásolhatja a T-rendszer működését.
Az immunrendszer szervei között megkülönböztetünk központi és perifériás szerveket. A központi szervek közé tartozik a csontvelő és a csecsemőmirigy, a perifériás szervek a lép és a nyirokcsomók. A csontvelőben a B-limfociták limfoid őssejtből, a T-limfociták limfoid őssejtből fejlődnek ki. Érésük során a T és B limfociták elhagyják a csontvelőt és a csecsemőmirigyet, és benépesítik a perifériás limfoid szerveket, megtelepedve a T és B zónában.
Miből áll a vér?
A vér formált elemekből (vagy vérsejtekből) és plazmából áll. A plazma a teljes vértérfogat 55-60%-át, a vérsejtek 40-45%-át teszik ki.
Vérplazma
A plazma enyhén sárgás, áttetsző folyadék, fajsúlya 1,020-1,028 (a vér fajsúlya 1,054-1,066), és vízből, szerves vegyületekből és szervetlen sókból áll. 90-92%-a víz, 7-8%-a fehérje, 0,1%-a glükóz és 0,9%-a sók.
Vérsejtek
vörös vérsejtek
A vörösvérsejtek vagy eritrociták a vérplazmában szuszpendálódnak. Számos emlős és ember vörösvérsejtjei magok nélküli, bikonkáv korongok. Az emberi vörösvértestek átmérője 7-8 µ, vastagsága 2-2,5 µ. A vörösvértestek képződése a vörös csontvelőben történik az érési folyamat során, elveszítik magjukat, majd bejutnak a vérbe. Egy vörösvérsejt átlagos élettartama körülbelül 127 nap, ezt követően a vörösvértestek elpusztulnak (főleg a lépben).
Hemoglobin
A lépben és a májban a régi vörösvértestekből származó hemoglobinmolekulák lebomlanak, a vasatomok újra felhasználódnak, a hem pedig lebontja és felszabadítja a májat bilirubinként és más epefestékként. Nukleáris vörösvérsejtek jelenhetnek meg a vérben nagy vérveszteség után, valamint akkor, ha a vörös csontvelő-szövet normál működése megzavarodik. Egy felnőtt férfi körülbelül 5 400 000 vörösvérsejtet tartalmaz 1 mm3 vérben, és egy felnőtt nő 4 500 000-5 000 000 vörösvértestet tartalmaz – 6-7 millió 1 mm3-ben. Minden vörösvérsejt körülbelül 265 millió hemoglobin molekulát tartalmaz, egy vörös pigmentet, amely oxigént és szén-dioxidot szállít. Becslések szerint másodpercenként körülbelül 2,5 millió vörösvérsejt termelődik, és ugyanennyi pusztul el. És mivel minden vörösvérsejt 265 · 106 molekula hemoglobint tartalmaz, körülbelül 650 · 1012 molekula ugyanabból a hemoglobinból képződik másodpercenként.
A hemoglobin két részből áll: fehérje - globin és vastartalmú - hem. A tüdő kapillárisaiban az oxigén a plazmából a vörösvérsejtekbe diffundál, és a hemoglobinnal (Hb) egyesülve oxihemoglobint (HbO2) képez: Hb + O2 «HbO2). A szöveti kapillárisokban alacsony parciális oxigénnyomás mellett a HbO2 komplex szétesik. Az oxigénnel kombinált hemoglobint oxihemoglobinnak, az oxigént leadott hemoglobint redukált hemoglobinnak nevezik. A CO2 egy része a vérben hemoglobinnal alkotott gyenge vegyület - karboxihemoglobin - formájában kerül szállításra.
Leukociták
A vér ötféle fehérvérsejtet vagy leukocitát tartalmaz, amelyek színtelen sejtmagot és citoplazmát tartalmaznak. A vörös csontvelőben, a nyirokcsomókban és a lépben képződnek. A leukocitákban nincs hemoglobin, és képesek aktív amőboid mozgásra. Kevesebb leukocita van, mint vörösvérsejt – átlagosan körülbelül 7000/1 mm3, de számuk 5000 és 9000 (vagy 10 000) között van különböző emberekben, sőt ugyanabban a személyben a nap különböző szakaszaiban: a legkevesebb korai életszakaszban. reggel, és leginkább délután. A leukociták három csoportra oszthatók: 1) szemcsés leukociták, vagy granulociták (citoplazmájuk granulátumokat tartalmaz), köztük vannak neutrofilek, eozinofilek és bazofilek; 2) nem szemcsés leukociták, vagy agranulociták, - limfociták; 3) monociták.
Vérlemezkék
Van egy másik csoportja a kialakult elemeknek - vérlemezkék vagy vérlemezkék, a legkisebbek az összes vérsejt közül. A csontvelőben képződnek. Számuk 1 mm3 vérben 300 000 és 400 000 között van. Fontos szerepet játszanak a véralvadási folyamat kezdetében. A legtöbb gerincesben
Az immunrendszer különbséget tesz „saját” és „nem-én” között, és elpusztítja a potenciálisan veszélyes idegen molekulákat és sejteket a szervezetből. Az immunrendszer képes azonosítani és elpusztítani saját szöveteinek patológiásan megváltozott sejtjeit. Az immunrendszer által felismert bármely molekula antigénnek (AG) számít.
A légutak és a gyomor-bél traktus bőre, szaruhártya és nyálkahártyája fizikai akadályt képez, amely az emberi szervezet első védelmi vonala. Ezen akadályok közül néhány aktív immunfunkcióval rendelkezik:
- Külső, keratinizált epidermisz: A bőr keratinocitái antimikrobiális peptideket (defenzineket), a faggyú- és faggyúmirigyek pedig csíragátló anyagokat választanak ki. Sok más immunsejt is jelen van a bőrben.
- A légzőrendszer, a gyomor-bélrendszer és a húgyúti nyálkahártya: a nyálkahártya antimikrobiális anyagokat tartalmaz, mint például lizozim, laktoferin és szekréciós immunglobulin A (SlgA).
Ha az immungátakat megsértik, 2 típusú immunitás valósul meg: veleszületett és szerzett. Számos molekuláris komponens játszik szerepet a veleszületett és szerzett immunitásban.
Veleszületett immunitás
A veleszületett (természetes) immunitás nem igényel előzetes antigénexpozíciót. Így azonnal válaszol az agresszornak. Főleg a széles körben bemutatott antigének molekuláit ismeri fel, nem pedig az adott szervezetre vagy sejtre specifikusakat. Összetevői a következők:
- fagocita sejtek
- antigénprezentáló sejtek
- természetes gyilkos sejtek,
- polimorfonukleáris leukociták.
A fagocita sejtek (vér neutrofilek és monociták, makrofágok és szöveti dendritikus sejtek) felszívják és elpusztítják a behatoló antigéneket. a kevésbé specifikus veleszületett védelmi rendszer részét képezik) opszonizálja AG. Az Ag-t prezentáló sejtek a lenyelt Ag fragmentumait mutatják be a T-limfocitáknak, és a szerzett immunitás részét képezik. A természetes gyilkos sejtek elpusztítják a vírussal fertőzött sejteket és egyes daganatsejteket.
Szerzett immunitás
A megszerzett immunitás megköveteli az antigénekkel való előzetes találkozást, pl. időre van szüksége a fejlődéshez az új agresszorral való kezdeti találkozás után. Gyors válasz következik. A rendszer megjegyzi a korábbi kapcsolatokat, és Ag-specifikus. Összetevői a következők:
- T-sejtek.
- A sejtekbe.
A bizonyos T-sejtes immunválaszokból származó szerzett immunitást sejt-közvetített immunitásnak nevezik. A B-sejtes reakciókból származó immunitást humorális immunitásnak nevezzük, mert. Az oldható Ag-specifikus antitestek kiválasztódnak a sejtekbe. A B-sejtek és a T-sejtek együtt dolgoznak az idegen elemek elpusztításában. Néhány ilyen sejtek nem pusztítják el közvetlenül az idegen anyagokat, hanem aktiválnak más fehérvérsejteket, amelyek felismerik és elpusztítják az idegen anyagokat.
Immunválasz
A sikeres immunvédelem megköveteli az immunválasz aktiválását, szabályozását és végrehajtását.
Aktiválás. Az immunrendszert az idegen Ag aktiválja, amelyet a keringő AT vagy sejtreceptorok ismernek fel. Ezek a receptorok lehetnek erősen vagy alacsony specifikusak. Az alacsony specifikus receptorok felismerik a mikroorganizmusok patogenitási faktorainak szerkezetében szereplő ligandumok közös csoportjait, mint például a Gram-negatív baktériumok lipopoliszacharidjai, a Gram-pozitív baktériumok peptidoglikánjai, a bakteriális flagellinek, a metilálatlan citozin-guanozin dinukleotidok (CpG motívumok) és a vírus kettős -szálas DNS. Aktiváció akkor is megtörténik, ha az AT-AG és a komplement-mikroorganizmus komplexek a sejtfelszíni receptorokhoz kötődnek az IgG Fc fragmentumához vagy a komplement C fragmentumokhoz.
A felismert antigén, antigén-antigén komplexek vagy komplement-mikroorganizmus fagocitózison megy keresztül. A legtöbb mikroorganizmust a fagocitózis elpusztítja, mások (például mikobaktériumok) elnyomják a fagociták azon képességét, hogy teljesen elpusztítsák őket, bár nem akadályozzák meg a felszívódást. Ilyen esetekben a T-limfociták által termelt citokinek, különösen az IgG, γ (IFN-γ) serkentik a lítikus enzimek és más mikrobicid anyagok termelését a fagociták által, amelyek elpusztítják a mikroorganizmusokat.
Míg az AG gyors fagocitózison megy keresztül és teljesen elpusztul (nem gyakori eset), a szerzett immunválasz működik. A keringő Ag-k esetében a lépből, a szöveti Ag-k esetében a nyirokcsomókból, a nyálkahártya-Ag-k esetében pedig a nyálkahártyához kapcsolódó limfoid szövetekből (pl. mandulák, adenoidok, Peyer-foltok) származik. Például a Langerhans dendritikus sejtek fagocitizálják az antigéneket a bőrben, és a helyi nyirokcsomókba vándorolnak, ahol az Ag-eredetű peptidek expresszálódnak a II. osztályú fő hisztokompatibilitási komplex (MHC) molekulák sejtfelszínén, amelyek bemutatják a peptidet a CD4 segítő sejtek (TH-k) számára. . Amikor egy T helper sejt kölcsönhatásba lép az MHC-peptid komplexszel, és kostimulációs jeleket kap, aktiválódik, és expresszálja az IL-2 citokin receptorait, és számos citokint szekretál. A T-helper sejtek mindegyike különböző anyagok kombinációit választja ki, ezáltal befolyásolja az immunválasz természetét.
Szabályozás. Az immunválaszt szabályozni kell, hogy elkerüljük a szervezetet érő szélsőséges károsodást (pl. anafilaxia, jelentős szövetkárosodás). A szabályozó T-sejtek segítenek az immunválasz szabályozásában az immunszuppresszív citokinek, például az IL-10 és a transzformáló növekedési faktor-β (TGF-β) szekrécióján, vagy egy rosszul ismert sejt-sejt kapcsolati mechanizmuson keresztül. Ezek a szabályozó sejtek megakadályozzák az autoimmun válasz kialakulását, és nyilvánvalóan hozzájárulnak a nem saját (idegen) Ag-kra adott válaszok megvalósításához.
Befejezés. Az immunválasz kiteljesedése akkor következik be, amikor az Ag-t leválasztják vagy eltávolítják a szervezetből. Antigén stimuláció nélkül a citokin szekréció leáll, és az aktivált citotoxikus sejtek apoptózison mennek keresztül. Az apoptózis a sejtet azonnali fagocitózisra jelöli, megelőzve a sejttartalom elvesztését és a gyulladás kialakulását. A memóriasejtekké differenciálódott T-sejtek és B-sejtek elkerülik ezt a sorsot.
Geriátriai komponens
Ahogy öregszünk, az immunrendszer kevésbé hatékony, nevezetesen:
- Gyengül az a képessége, hogy felismerje a sajátját az idegentől, így nő az autoimmun betegségek gyakorisága.
- A makrofágok kevésbé intenzíven pusztítják el a baktériumokat, rákos sejteket és más antigéneket, ami megmagyarázza az idősek körében tapasztalható rákos megbetegedések növekedését.
- A T-sejtek nem képesek gyorsan reagálni az Ag-re.
- Csökken az új Ag-re reagálni képes limfociták száma.
- Az öregedő szervezet kevesebb komplementet termel bakteriális fertőzésekre válaszul.
- Kevesebb AT termelődik a magas vérnyomásra válaszul, és az AT-k kevésbé képesek kötődni a magas vérnyomáshoz, ami megmagyarázza a tüdőgyulladás, influenza, fertőző endocarditis és tetanusz megnövekedett előfordulási gyakoriságát, valamint az e patológiák miatti halálozási kockázat növekedését az időseknél. Ezek a változások részben magyarázhatják az idősek körében végzett védőoltások hatékonyságának hiányát is.
Az immunrendszer összetevői
Az immunrendszer sejt- és molekuláris komponensekből áll, amelyek együtt működnek az Ag-ek elpusztításában.
Antigén prezentáló sejtek
Bár egyes antigének közvetlenül stimulálhatnak immunválaszt, a T-függő szerzett immunválaszhoz általában antigénprezentáló sejtek (APC) jelenlétére van szükség, amelyek antigénpeptideket mutatnak be MHC-molekulákkal komplexben. Az intracelluláris Ag-k (például a vírusok) bármely sejtmaggal rendelkező sejt által átalakíthatók és bemutathatók a Tc limfociták CD8 receptoraivá. Az ezeket kódoló fehérjék segítségével, amelyek megzavarják ezt a folyamatot, egyes vírusok (például citomegalovírus) elkerülik a pusztulást. Az intracelluláris Ag-t peptiddé kell alakítani, és MHC II. osztályú molekulákkal komplexben kell bemutatni az APC-k felületén, hogy a CD4 sejteket hordozó helper sejtek felismerjék.
A véráramban lévő monociták a szöveti makrofágok előfutárai. A monociták a szövetekbe vándorolnak, ahol 8 óra elteltével makrofágokká fejlődnek a különböző sejttípusok (pl. endoteliális sejtek, fibroblaszt sejtek) által kiválasztott makrofág telep-stimuláló faktor (M-CSF) hatására.
A makrofágokat az IFN-γ és a granulocita-makrofág telep-stimuláló faktor (GM-CSF) aktiválja. Az aktivált makrofágok elpusztítják az intracelluláris organizmusokat, és IL-1-et és tumor nekrózis faktor-alfát (TNF-α) választanak ki. Ezek a citokinek potencírozzák az IFN-γ és a GM-CSF szekrécióját, és fokozzák az adhéziós molekulák expresszióját az endothelsejtek felszínén, megkönnyítve a leukociták beáramlását a fertőzés helyére és a patogén faktor elpusztítását. A makrofágokat génexpressziós profiljuk alapján altípusokba sorolták.
A dendritikus sejtek a bőrben (Langerhans-sejtek), a nyirokcsomókban és az egész test szöveteiben jelen vannak. A bőrben található dendritikus sejtek határvonali APC-k, amelyek befogják az antigént és eljuttatják azt a helyi nyirokcsomókba, ahol aktiválják a T-limfocitákat.
Mindazonáltal rendelkeznek receptorokkal az IgG és a komplement Fc-fragmensére, ami lehetővé teszi számukra, hogy immunkomplexeket kötődjenek, és bemutassák azokat a másodlagos limfoid szervek csíraközpontjainak B-limfocitáihoz.
Polimorfonukleáris leukociták
A polimorfonukleáris leukocitákat (PMN) granulocitáknak is nevezik, mert. citoplazmájuk specifikus szemcséket tartalmaz.
A keringő vérben jelen vannak, és szegmentált sejtmagjuk van, kivéve a hízósejteket, amelyek folyamatosan jelen vannak a szövetekben, és funkcionálisan hasonlóak a keringő bazofilekhez.
A neutrofilek az összes leukocita 40-70%-át teszik ki; ők jelentik az első védelmi vonalat a fertőzések elleni küzdelemben. Az érett neutrofilek felezési ideje 2-3 nap. Akut gyulladásos folyamat (például fertőző) során a kemotaktikus tényezőkre reagálva a neutrofilek elhagyják a véráramot és bejutnak a szövetekbe. Céljuk a patogén faktorok fagocitózisa és elpusztítása. A mikroorganizmusok elpusztulnak, amikor a fagociták lítikus enzimeket és reaktív oxigénfajtákat (szuperoxid, hipoklórsav) termelnek, vagy granulátumtartalom (defenzinek, proteázok, szöveti permeabilitást növelő baktericid fehérjék, laktoferrin és lizozim) felszabadulását idézik elő. DNS és hisztonok is felszabadulnak, és ezek a szemcsetartalommal, például az elasztázzal együtt rostokat képeznek a környező szövetekben, amelyek segíthetnek a baktériumok elpusztításában és az enzimaktivitás lokalizálásában.
A bazofilek a fehérvérsejtek kevesebb mint 5%-át teszik ki, és hasonlóak a hízósejtekhez, bár különböző sejtvonalakhoz tartoznak. Mindkét sejt nagy affinitású IgE receptorral rendelkezik. Amikor ezek a sejtek specifikus antigénnel találkoznak, ez az antigén keresztkötéseket köt a szomszédos bivalens IgE molekulákkal, ami sejtdegranulációt okoz, kész gyulladásos mediátorok felszabadulásával és új mediátorok (leukotriének, prosztaglandinok, tromboxánok) képződésével.
A hízósejtek a test különböző szöveteiben találhatók. A nyálkahártya hízósejtjeiben a szemcsék triptázt és kondroitin-szulfátot tartalmaznak, ha pedig a sejt kötőszövetben lokalizálódik, akkor a szemcséi triptázt, kimázt és heparint tartalmaznak. Amikor ezek a mediátorok felszabadulnak, protektív akut gyulladásos válasz alakul ki. A degranulációt az anafilatoxin, a C3a és C5a komplement fragmentumok válthatják ki.
Citotoxikus leukociták
A citotoxikus leukociták közé tartoznak:
- Természetes gyilkos sejtek.
- Limfokin-aktivált gyilkos sejtek.
Természetes gyilkos sejtek (NKC). A tipikus NK-sejtek a perifériás vér mononukleáris sejtjeinek 5-15%-át teszik ki. Kerek sejtmaggal és szemcsés citoplazmával rendelkeznek. Az NK-sejtek apoptózist indukálnak a fertőzött és abnormális sejtekben különböző utakon keresztül. Veleszületett válaszsejtekként nem rendelkeznek antigén-specifikus receptorokkal és immunológiai memóriával.
A tipikus ECC-k nagyon fontosak a mutáló sejtek szabályozásában, mert aktiváló és gátló receptorokat egyaránt expresszálnak. Az aktiváló NK-receptorok számos célsejt-ligandumot felismernek (például MHC I. osztályú A lánc és B lánc. A gátló NK receptorok az MHC I. osztályú molekulákat ismerik fel. Az NK csak akkor pusztítja el célpontjaikat, ha nincs erős jel a gátló receptoroktól. A jelenléte MHC I. osztályú molekulák (normálisan sejtmagos sejteken expresszálódó) sejteken való elpusztítása megakadályozza, hogy a sejt elpusztuljon, és a hiány azt jelzi, hogy a sejt valamilyen vírussal fertőzött, amely gátolja az MHC expresszióját, vagy elvesztette az MHC expresszióját, mert a rák megváltoztatta a sejtet. Az NK-hiányban szenvedő betegek különösen érzékenyek a herpesz fertőzésre és a humán papillomavírusra.
Az NK-sejtek számos citokint is szekretálnak; ezek az IFN-γ fő forrásai. Az IFN-γ kiválasztásával az NK-sejtek befolyásolhatják a szerzett immunrendszert az 1-es típusú (Tn1) helper sejtek differenciálódásának (differenciálódásának) elősegítésével és a 2-es típusú (Tn2) helper sejtek gátlásával.
Limfokin-aktivált gyilkos (LAK) sejtek. Egyes limfociták nagyon erős limfokin-aktivált ölősejtekké (LAK) fejlődnek, amelyek a tumorsejtek és a kóros limfociták széles skáláját képesek elpusztítani (például bizonyos vírusokkal fertőzötteket). Ezek a sejtek nemcsak a limfociták egyedi sejtaltípusát alkotják, hanem fenomenálisak is. A LAK prekurzorok heterogének, de kezdetben NK-szerű sejtek (leggyakrabban) vagy T-limfocitaszerű sejtek közé sorolhatók.
Limfociták
A limfociták két legfontosabb típusa:
- B-limfociták, amelyek a csontvelőben érnek.
- T-limfociták, amelyek a csecsemőmirigyben érnek.
Morfológiájukban nem különböznek egymástól, de eltérő immunfunkciókkal rendelkeznek. Ezeket az Ag-specifikus felszíni receptorok, a differenciálódási klasztereknek (CD-k) nevezett molekulák különböztetik meg egymástól, amelyek egy adott sejtaltípusban jelen vannak vagy hiányoznak. Több mint 300 CD azonosított. Minden limfocita egy specifikus antigént ismer fel a felszíni receptorokon keresztül.
B limfociták. A vér limfocitáinak 5-15%-a B-limfocita. Jelen vannak a lépben, a nyirokcsomókban a nyirokszövetek nyálkahártyáján is. A B-sejtek Ag-t mutathatnak be a T-sejteknek, de elsődleges funkciójuk az, hogy plazmasejtekké fejlődjenek, amelyek antitesteket (AT) termelnek és választanak ki. A B-sejtes immunhiányban (pl. X-hez kötött agammaglobulinémia) szenvedő betegek különösen érzékenyek a visszatérő bakteriális fertőzésekre.
Az Ig-t kódoló gének véletlenszerű átrendeződését követően a B-limfociták szinte végtelen számú egyedi Ag-t képesek felismerni. A génátrendeződés szekvenciálisan megy végbe a B-sejtek fejlődése során a vörös csontvelőben. A folyamat egy elkötelezett őssejttel kezdődik, átmegy a pro-B és pre-B limfociták stádiumain, és éretlen B-limfocitával ér véget. Ha ez az éretlen B-limfocita kölcsönhatásba lép az Ag-vel, akkor ennek a sejtnek inaktiválása (tolerancia kialakulása) vagy eliminációja (apoptózisa) történhet. Az éretlen B-limfociták, amelyek nem mentek át inaktiváláson vagy elimináción, tovább fejlődhetnek az érett fiatal B-limfociták állapotába, elhagyják a vörös csontvelőt, és a perifériás limfoid szervekbe költözhetnek, ahol magas vérnyomással találkozhatnak. A magas vérnyomásra adott válaszuk két szakaszban történik:
- Elsődleges immunválasz. Amikor az érett fiatal B-limfociták először találkoznak Ag-vel, ezek a sejtek blast transzformáción, klonális proliferáción és memóriasejtekké differenciálódnak, amelyek a jövőben ugyanarra az Ag-re reagálnak, vagy érett AT-termelő plazmasejtekké. Az AT gyártása előtt több napos látenciaidő áll rendelkezésre. Ekkor már csak IgM termelődik. Kezdetben csak IgM termelődik. A T-limfocitákkal való interakciót követően a B-limfocitákban az Ig gének további átrendeződése következhet be, ami átváltja a szintézist IgG-re, IgA-ra vagy IgE-re.
- Másodlagos immunválasz (anamnesztikus, fokozott). Amikor a memória B-sejtek és a T-helper sejtek újra találkoznak ugyanazzal az Ag-vel. A memória B-sejtek gyorsan szaporodnak, érett plazmasejtekké differenciálódnak, gyorsan szintetizálnak és nagy mennyiségű AT-t (főleg IgG-t, mivel a T-limfociták ennek az izotípusnak a szintézisét váltanak ki) a vérbe és más szövetekbe, ahol az AT reakcióba léphet Ag . Így a magas vérnyomással való második találkozás után az immunválasz gyorsabb és hatékonyabb. T limfociták.
A T-limfocitáknak 3 fő típusa van:
- Segítő.
- Szabályozó.
- Citotoxikus.
Az érettebb T-limfociták CD4-et vagy CD8-at, valamint antigénkötő Ig-szerű receptorokat, úgynevezett T-sejt-receptorokat (TCR-ek) expresszálnak. A TCR-t kódoló gének az immunglobulin génekhez hasonlóan átrendeződnek. Ennek eredményeként bizonyos specifitás és affinitás érhető el az APC membránon prezentált és AG peptidekhez kapcsolódó MHC-molekulákkal való érintkezéskor. A T-limfocitákban a specifikus kapcsolatok száma szinte végtelen.
A T-limfocita aktiválásához a TCR vagy az Ag-MHC komplexhez vagy a segédmolekulákhoz kötődik; ellenkező esetben a T-limfocita inaktiválva marad, vagy apoptózissal elpusztul. Egyes segédmolekulák gátolják a korábban aktivált T-limfocitákat, és így leállítják az immunválaszt. A CTLA-4 gén polimorfizmusa bizonyos autoimmun patológiákkal jár.
A T helper (Tn) sejtek általában CD4, de lehetnek CD8 is. A Th0 sejtektől a következők egyikére különböznek:
Minden sejttípus specifikus citokineket választ ki. A citokintermelésnek különböző általános mintái vannak, amelyek meghatározzák a Tn-sejtek funkcionális fenotípusait. A Tn2 sejtek képesek egymás funkcionális aktivitását egy bizonyos szintig leszabályozni, ami a Tn1 vagy Tn2 válasz dominanciájához vezet.
A különböző típusú β-sejtek közötti különbség klinikailag jelentős. Például a Tn1 válasz dominál a tuberkuloid leprában, és a Tn2 válasz a leprátás leprában. A Tn1 válasz bizonyos autoimmun kórképekre jellemző, a Tn2 válasz pedig elősegíti az IgG termelődését és az allergiás betegségek kialakulását, valamint egyes autoimmun patológiákban (például Graves-kór, myasthenia gravis) segíti a B-sejtek antitestek kiválasztását. Az immunhiányos betegeket hibás Tn 17 sejtek jellemzik (például hyper IgE szindróma), ezek a betegek a leginkább érzékenyek a Candida albicans és a Staphylococcus aureus által okozott fertőzésekre.
Szabályozó T-sejtek. Közvetítik az immunválaszok elnyomását, és jellemzően a Fox3 transzkripciós faktort fejezik ki. Ez a folyamat a CD4 CD8 professzionális sejtalcsaládokat érinti, amelyek vagy immunszuppresszív tulajdonságokkal rendelkező citokineket választanak ki, vagy elnyomják az immunválaszt, és a szuppresszió mechanizmusa még mindig kevéssé ismert, és közvetlen érintkezést igényel a sejtek között. A Foxp3 funkcionális mutációival rendelkező betegeknél autoimmun patológia, IPEX-szindróma alakul ki (immunreguláció, poliendokrinopátia, enteropathia, X-hez kötött).
A citotoxikus T(Tc) sejtek általában CD8, de lehetnek CD4 is; szükségesek az intracelluláris kórokozók, különösen a vírusok elpusztításához.
A Tc-sejtek 3 fejlődési szakaszon mennek keresztül:
- Progenitor sejt, amely megfelelő stimuláció esetén Tc sejtté differenciálódik.
- Egy differenciált effektor sejt, amely képes elpusztítani a célpontokat.
- Nyugalomban lévő (már nem stimulált), de az antigének és MHC eredeti kombinációjával történő ismételt stimuláció után készen áll az effektor funkció végrehajtására.
A teljesen aktivált Tc-sejtek az NK-sejtekhez hasonlóan képesek megölni a fertőzött célsejtet azáltal, hogy apoptózist indukálnak.
A TC sejtek lehetnek:
- Izogén: vírusfertőzés vagy más idegen fehérje által módosított saját (autológ) sejtekre válaszul termelődik.
- Allogén: idegen MHC-termékeket expresszáló sejtekre reagálva termelődik (például szervátültetések során, amikor a donor MHC-molekulák eltérnek a recipiens MHC-től) Egyes Tc-sejtek specifikusan felismerik az idegen MHC-t (közvetlen útvonal); mások felismerhetik a transzplantált recipiens saját MHC-molekulái által bemutatott idegen MHC-fragmenseket (indirekt útvonal).
Antitestek
Az AT antigénreceptorként működik a B-sejtek felszínén, és a plazmasejtek választják ki az Ag-re adott válaszként. Az AT-k specifikus konfigurációkat ismernek fel az Ag-ek felszínén (pl. fehérjék, poliszacharidok, nukleinsavak). Az AT és az AG pontos egyezést jelent, mert alakjuk és egyéb felületi tulajdonságaik (például terhelés) kiegészítik egymást. Ugyanaz az AT-molekula keresztreakcióba léphet a megfelelő Ag-vel, ha eptopjai kellően hasonlóak az eredeti Ag epitópjaihoz.
Szerkezet. Az AT-k 4 polipeptid láncból állnak (2 azonos nehéz lánc és 2 azonos könnyű lánc), amelyeket diszulfid kötések kapcsolnak össze, hogy létrehozzák az Y konfigurációt. Mind a nehéz, mind a könnyű láncnak van egy változó (V) és egy állandó (C) régiója.
V - A variábilis régiók Y felső részének aminoterminális végein helyezkednek el; változónak nevezik őket, mert különböző aminosavakat tartalmaznak, amelyek meghatározzák az lg specificitását. A hipervariábilis régiók idiotípusos determinánsokat tartalmaznak, amelyekhez bizonyos természetes (anti-idiotípusos) AT-k kötődnek; ez a kapcsolat segíthet a B humorális válasz szabályozásában. A B-limfociták képesek megváltoztatni a termelt Ig nehéz lánc izotípusát, de megtartják a V-régió nehéz láncait és a teljes könnyű láncot, és így megtartják az antigénspecifitást.
A C régió viszonylag állandó aminosavszekvenciából áll, amely minden IgG izotípusra jellemző.
Az AT amino-terminális (variábilis) vége kötődik az Ag-hez, és AGAT komplexet képez. Az lg Ag-kötő része (Fab) egy könnyű láncból és egy nehéz lánc fragmentumból áll, és tartalmazza az lg molekula V régióját (vegyes rész).
Antitest osztályok. Az antitestek 5 osztályba sorolhatók:
Ezek az osztályok különböznek a nehéz lánc típusától; Kétféle könnyű lánc is létezik (k és A). Mind az 5 lg osztálynak van k- vagy λ-könnyű lánca.
Az IgM az első AT, amely új AG találkozása után képződik. 5 Y molekulából áll (10 nehéz és 10 könnyű lánc), amelyeket egyetlen kötéssel kapcsolnak össze. Az IgM túlnyomórészt az intravaszkuláris térben kering; megköti és agglutinálja az Ag-t, és aktiválhatja a komplementet, ami elősegíti a fagocitózist. Az IgM izohemagglutininek és számos Gram-negatív mikroorganizmus AT-je. Az IgM monomer a B-limfociták felszínén található antigénreceptor. A hiper-IgM-szindrómában szenvedő betegeknél az antitestek egy specifikus osztályának (például CD40-et vagy CD154-et kódoló gének) beépítésében szerepet játszó gének hibái vannak; ezért IgA-, IgM- és IgE-szintjük alacsony vagy hiányzik, a keringő IgM-szintek pedig gyakran magasak.
Az IgG az IgG domináns izotípusa; intra- és extravascularis térben egyaránt kering. Az IgG az elsődleges keringő IgG, amely az újraimmunizálás után (másodlagos immunválasz során) jelenik meg, és a domináns izotípus a kereskedelmi forgalomban kapható globulintermékekben. Az IgG megvédi a szervezetet a baktériumoktól, vírusoktól, méreganyagoktól, és ez az egyetlen Ig izotípus, amely átjut a placenta gáton. Ezért fontos az újszülöttek védelmezőjeként az antitestek ezen osztálya, de a patogén IgG antitestek, ha jelen vannak a kismama szervezetében, súlyos kóros magzati állapotot válthatnak ki.
Az IgG-nek 4 alosztálya van: IgG1, LgG2, IgG3, IgG4, amelyek számozása az IgG szérumkoncentrációi szerint csökkenő sorrendben történik. Az IgG alosztályok elsősorban komplement aktiváló képességükben különböznek egymástól; Az IgG1 és az LgG3 a leghatékonyabb, az IgG2 kevésbé hatékony és az LgG4 hatástalan. Az IgG1 és IgG3 hatékony mediátorai az antitestek által közvetített sejtes citotoxicitásnak; Az IgG4 és az IgG2 kevésbé hatékony ebben a tekintetben.
Az IgA jelen van a nyálkahártyák felületén, a szérumban és a váladékokban (nyál, könnyfolyadék, légúti, gyomor-bél- és húgyúti váladék, kolosztrum), ahol kezdeti antibakteriális és vírusellenes védelmet nyújt. A J lánc az IgA-t dimerré köti – szekréciós IgA molekula keletkezik. A szekréciós IgA-t a plazmaciták szintetizálják a gyomor-bél traktus és a légutak nyálkahártyájának szubepitheliális részében. A szelektív IgA-hiány viszonylag gyakori, de klinikai jelentősége csekély, mivel az antitestek más osztályai között keresztfunkcionalitás létezik.
Az IgD az IgM-mel együtt expresszálódik a fiatal B-limfociták felszínén. Még mindig nem világos, hogy ennek a 2 osztálynak különböző funkciói vannak-e, és ha igen, milyen mértékben. Lehetnek egyszerűen a molekuláris degradáció példái. A szérum IgD szintje nagyon alacsony, és a keringő IgD funkciója nem ismert.
Akut fázisú reagensek
Az akut fázisú reagensek plazmafehérjék, amelyek szintje élesen megemelkedik, vagy bizonyos esetekben csökken a fertőző folyamatok vagy szövetkárosodás során. A legjelentősebb növekedés a C-reaktív fehérje és a mannóz-kötő lektin (amely rögzíti a komplementfehérjéket és egy opszonin szerepet játszik), az α 1-savas glikoprotein transzportfehérje és a szérum amiloid komponens CRP és ESR gyakran mérhető; az emelkedett szint a fertőzés vagy gyulladás nem specifikus jele. A megnövekedett ESR fő oka a megnövekedett fibrinogén.
Sok akut fázisú reagens termelődik a májban. Együtt segítik a szövetkárosodás korlátozását, növelik a szervezet fertőzésekkel szembeni ellenálló képességét, elősegítik a szövetek helyreállítását és megállítják a gyulladást.
Citokinek
A citokinek olyan polipeptidek, amelyeket immun- és más sejtek választanak ki specifikus antigénekkel, endotoxinnal és más citokinekkel való kölcsönhatásuk után. A citokinek fő csoportjai az interferonok:
- interferonok;
- tumornekrózis faktorok (TNF-α, limfotoxinok-α, limfotoxinok-β);
- interleukinok (IL);
- kemokinek;
- átalakuló növekedési faktorok;
- hematopoietikus kolónia-stimuláló faktorok (CSF).
Bár a limfociták egy specifikus antigénnel való kölcsönhatás után citokint kezdenek kiválasztani, maguk a citokinek nem antigén-specifikusak.
A citokinek jeleket továbbítanak a sejtfelszíni receptorokon keresztül. Például az I/1-2 receptor 3 láncból áll: α, β és γ. Az IL-2 receptor affinitása magas lesz, ha mind a 3 lánc expresszálódik, közepes, ha csak a β és y lánc expresszálódik, és alacsony, ha csak az α lánc expresszálódik. A lánc mutációi vagy deléciói képezik az X-hez kötött súlyos kombinált immunhiány alapját.
A kemokinek kemotaxist és leukociták migrációt váltanak ki. 4 alcsalád létezik, amelyek az első két ciszteincsoport között elválasztó aminosavak számában különböznek. A kemokin receptorok (CCR5 a memória T-limfocitákon, monociták/makrofágok, dendritikus sejtek; CXCR4 más T-limfocitákon) a HIV (humán immundeficiencia vírus) sejtbe való bejutásának koreceptorai.
Humán leukocita antigének
A humán leukocita antigén (HLA) rendszer a 6. kromoszómán lokalizálódik. Ez a kromoszóma sejtfelszíni molekulákat kódol.
Az I. osztályú MHC molekulák minden sejtmag felszínén jelen vannak transzmembrán glikoproteinként; Miután ezek a molekulák denaturálódnak és lebomlanak, a vérlemezkék felveszik őket. Egy normál I. osztályú molekula nehéz láncból áll, amely egy p2 mikroglobulin molekulához kapcsolódik. A nehézlánc két kapcsolódó peptiddoménből, egy Ig-szerű doménből, egy transzmembrán régióból és egy citoplazmatikus végből áll. Az MHC I. osztályú molekula nehézláncát a HLA-A, -B vagy C lókuszok génjei kódolják. Az MHC I. osztályú molekulákra reagáló limfociták CD8 molekulákat expresszálnak és effektor funkciókat látnak el, amelyek közé tartozik a fertőzött sejtek felismerésének képessége. Mivel minden sejtmaggal rendelkező sejt MHC I. osztályú molekulákat expresszál, minden fertőzött sejt antigénprezentáló a CD8 pozitív T limfociták számára (a CD8 a nem polimorf I. osztályú nehézlánc régióhoz kötődik). Egyes MHC I. osztályú gének nem klasszikus MHC-molekulákat kódolnak, például HLA-G-t és HLA-E-t (amelyek peptideket mutatnak be egyes NK-receptoroknak).
Az MHC II. osztályú molekulák jellemzően csak a professzionális Ag-t prezentáló sejteken, a csecsemőmirigy-hámsejteken és az aktivált (de nem nyugvó) T-sejteken vannak jelen; A legtöbb sejtmaggal rendelkező sejt interferon (IFN)-γ segítségével stimulálható MHC II. osztályú molekulák expressziójára. Az MHC I. osztályú molekulák két polipeptid (a és (3) láncból állnak; mindegyik peptidnek van egy peptidkötő régiója, egy lg-szerű régiója és egy transzmembrán régiója citoplazmatikus farokkal. Mindkét polipeptid láncot a HLA- gének kódolják DP, -DQ vagy -DR kromoszóma 6. Az MHC II. osztályú molekulákra reagáló limfociták CD4-et expresszálnak, és gyakran segítő T-sejtek.
Az MHC III. osztályú régió számos gyulladásban fontos molekulát kódol.
A szerológiai tipizálással azonosított egyedi antigének, amelyeket az I. és II. osztályú lókuszok génjei kódolnak, szabványos elnevezéssel rendelkeznek. A DNS-szekvenálással meghatározott allélok tartalmazzák a gén nevét, amelyet egy csillag követ, majd az allélcsoportot jelző számok (gyakran az allél által kódolt szerológiailag azonosított antigénnek felelnek meg), majd egy kettőspont és az allélt jelző számok. Néha az alléljelölésben további számok szerepelnek a kettőspont után az azonos fehérjéket kódoló allélvariánsok jelölésére, a második kettőspont után pedig számokat adunk az intronokban vagy az 5" vagy 3"-os nem lefordított régiókban lévő polimorfizmusok jelzésére.
Az MHC I. és II. osztályú molekulák a leginkább immunogén antigének, amelyeket az allogén transzplantátum kilökődés során ismernek fel. A legerősebb determináns a HLA-DR, ezt követi a HLA-B és az -A, ezért ez a három lókusz a legfontosabb a recipiens számára megfelelő (szövetkompatibilis) donor kiválasztásánál.
Kiegészítő rendszer
A komplementrendszer olyan enzimek kaszkádja, amelyek elősegítik a fertőző folyamat elleni küzdelmet. Ez a rendszer összekapcsolja a veleszületett és szerzett immunitást:
- Növeli az antitest (AT) választ és az immunológiai memóriát.
- Idegen molekulák bérbeadása.
- Az immunkomplexek eltávolítása. A komplementrendszer komponensei számos biológiai funkciót látnak el.
Komplement fehérjék aktiválása: a komplement aktiválásának 3 módja van:
- klasszikus,
- lektin (mannózkötő lektin-MBL),
- alternatív.
A klasszikus útvonal összetevőit C betű és egy szám jelöli, amely jelzi az azonosítás sorrendjét. Az alternatív útvonal komponenseit gyakran betűvel (pl. B faktor, D faktor) vagy külön névvel (pl.propindin) jelölik.
A klasszikus módszer. A klasszikus útvonal aktiválása egy AT-függő folyamat, amely a C1 és az Ag-lgM vagy Ag-lgG komplexekkel való kölcsönhatása után kezdődik, vagy egy AT-független folyamat, amikor polianionok (heparin, protamin, apoptotikus sejtek DNS-e vagy RNS-e) Gram-negatív baktériumok vagy kötött C-reaktív fehérje közvetlenül reagál a C1-gyel. Ezt az utat a C1 inhibitor (C1-INM) szabályozza. Az örökletes angioödéma a C1-INH genetikai hiányával jár.
A lektin út (mannóz-kötő lektin) egy AT-független folyamat; akkor kezdődik, amikor az MBL tejsavófehérje mannózhoz, fruktózhoz kötődik.
Egy alternatív útvonal a mikrobiális sejtfelszíni komponensek vagy lg kis mennyiségű C3-hoz való adhéziójával kezdődik. Ezt az utat a megfelelő H faktor, a nekrózist gyorsító faktor szabályozza.
Ez a 3 útvonal végül konvergál, amikor a C3 konvertáz a C3-at C3a-vá és C3b-vé alakítja. A C3 hasítása a membrán támadó komplex (MAC) kialakulásához vezethet, amely a komplement rendszer citotoxikus komponense. A MAC az idegen sejtek lízisét okozza.
A komplement komponens hiányában szenvedő betegek gyakran érzékenyek a visszatérő bakteriális fertőzésekre, különösen a C3 komponens hiányában. A C1 és C4 hibái a szisztémás lupus erythematosushoz kapcsolódnak.
Biológiai aktivitás. A komplementrendszer komponensei más biológiai funkciókat is ellátnak, amelyeket a komplementreceptorok (CR-ek) hajtanak végre különböző sejttípusokon.
- A CR1 (CD35) elősegíti a fagocitózist, és részt vesz az immunkomplexek kiürülésében.
- A CR2 (CD21) szabályozza az AT B-limfociták termelését, és az Epstein-Barr vírus receptora.
- A CR3 (CDllb/CD18), SR4 (CDllc/CD18) és Clq receptorok szerepet játszanak a fagocitózisban.
- A C3a, C5a és C4a (gyengén) anafilatikus aktivitást mutatnak. A hízósejtek degranulációját okozzák, ami fokozott érpermeabilitáshoz és simaizom-összehúzódáshoz vezet.
- A C3b opszoninként működik, bevonja a mikroorganizmusokat, és ezáltal fokozza fagocitózisukat.
- A C3d fokozza a B-limfociták AT termelését.
- A C5a egy neutrofil kemoettraktáns Szabályozza a neutrofilek és a monociták aktivitását, és fokozott sejtaggregációt, degranulációt és intracelluláris enzimek felszabadulását okozhatja a granulocitákból, toxikus oxigén metabolitok termelődését és egyéb, a sejtmetabolizmushoz kapcsolódó hatásokat.
A fő sejtes immunkomponensek közé tartozik az összes vérleukocita, amely az ún immunkompetens sejtek. Az érett leukociták öt sejtpopulációt kombinálnak:
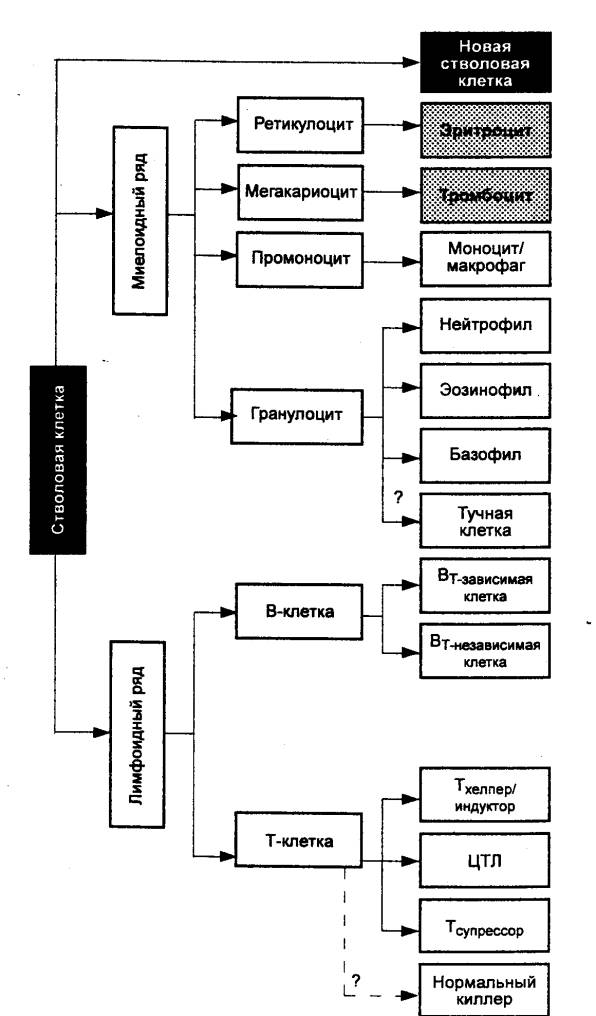
limfociták, monociták, neutrofilek, eozinofilek és bazofilek. Immunkompetens sejtek a test szinte bármely részében megtalálhatók, de elsősorban a kialakulásuk helyén - az elsődleges és másodlagos nyirokszervekben - koncentrálódnak (8.1. ábra). Ezen sejtek kialakulásának elsődleges helye a hematopoietikus szerv - vörös csontvelő melynek melléküregeiben monociták és valamennyi granulocita (neutrofilek, eozinofilek, bazofilek) képződnek és teljes differenciálódási cikluson mennek keresztül. Itt kezdődik a limfociták differenciálódása. Az összes populáció leukocitái egyetlen csontvelőből származnak, pluripotens vérképző őssejt, melynek medencéje önfenntartó (8.2. ábra).
Az őssejtek differenciálódásának különböző irányait a csontvelői vérképzés gócaiban lévő specifikus mikrokörnyezetük és a specifikus hematopoietikus faktorok, köztük a telepstimuláló faktorok, kelonok, prosztaglandinok és mások termelése határozza meg. A csontvelőben az immunkompetens sejtek képződését és differenciálódását irányító rendszer ezen tényezőkön túlmenően a szervezet egészére kiterjedő szabályozó anyagok egy csoportját foglalja magában, amelyek közül a legfontosabbak a hormonok és az idegrendszer mediátorai.
A szervezetben a limfocitákat két nagy alpopuláció képviseli, amelyek hisztogenezisében és immunfunkcióiban különböznek egymástól. Ez T limfociták, sejtes immunitás biztosítása, és B limfociták, felelős valamiért
osu antitestképződés, azaz humorális immunitás létrehozása. Ha a B-limfociták a csontvelőben a teljes differenciálódási cikluson átesnek érett B-sejtekké, akkor a T-limfociták a pre-T-limfociták stádiumában a véráramon keresztül egy másik elsődleges limfoid szervbe - a csecsemőmirigybe - vándorolnak. differenciálódásuk az érett T-sejtek összes sejtformájának kialakulásával ér véget.
Alapvetően különbözik tőlük a limfociták egy speciális alpopulációja - normál (természetes) gyilkos sejtek(NK) és K-sejtek. Az NK-k olyan citotoxikus sejtek, amelyek előzetes immunizálás nélkül, azaz antitestek hiányában pusztítják el a célsejteket (főleg a tumorsejteket és a vírussal fertőzött sejteket). A K-sejtek képesek elpusztítani a kis mennyiségű antitesttel bevont célsejteket.
Az érést követően az immunkompetens sejtek bejutnak a véráramba, amelyen keresztül a monociták és a granulociták a szövetekbe vándorolnak, a limfociták pedig a másodlagos limfoid szervekbe kerülnek, ahol differenciálódásuk antigénfüggő fázisa következik be. A keringési rendszer az immunkomponensek, köztük az immunkompetens sejtek szállításának és újrahasznosításának fő útvonala. A vérben általában nem fordulnak elő immunológiai reakciók. A véráramlás csak a sejteket a működési helyükre szállítja.
Granulociták(neutrofilek, eozinofilek, bazofilek) a csontvelőben történő érés után csak effektor funkciót látnak el, ami után egyszer elpusztulnak. Monociták a csontvelőben történő érés után szövetekben telepednek meg, ahol a belőlük képződött szöveti makrofágok is effektor funkciót látnak el, de hosszan és ismétlődően. Minden más sejttől eltérően, limfociták a csontvelőben (B-sejtek) vagy a csecsemőmirigyben (T-sejtek) történő érés után a másodlagos limfoid szervekbe jutnak (8.3. ábra), ahol
|
Rizs. 8.1 Lymphomyeloid komplexum BM - csontvelő; KS - erek; LTK - bél limfoid szövet; LS - nyirokerek; LU - nyirokcsomók; SL - lép; T - csecsemőmirigy (csecsemőmirigy). |
|
|
|
|
|
Rizs. 8.2 Multipotens hematopoietikus őssejtés leszármazottai CTL - citotoxikus T-limfocita (T-killer). |
|
fő funkciójuk a szaporodás egy antigén inger hatására rövid életű specifikus effektor sejtek és hosszú életű memóriasejtek megjelenésével. "Immunológiai memória - a szervezet azon képessége, hogy egy antigén ismételt beadására olyan immunválaszt tudjon reagálni, amelyet az első immunizálásnál erősebb és gyorsabb válasz jellemez.
Másodlagos limfoid szervek szétszórva a testben, hogy kiszolgálja az összes szövetet és felületet. A másodlagos limfoid szervek közé tartozik a lép, a nyirokcsomók, a nyálkahártyák közelében lévő limfoid szövetek - a vermiform függelék (függelék), a Peyer-foltok, a mandulák és a garat limfoid gyűrű egyéb képződményei, magányos (egyetlen limfoid tüszők). a bélben és a hüvelyben, valamint a limfoid sejtek diffúz felhalmozódása a test összes nyálkahártyájának szubepitheliális tereiben, valamint az újonnan kialakult limfoid szöveti gócok a granulációs szövetben a krónikus gyulladásgócok körül.
A másodlagos limfoid szervekben a T- és B-limfociták először érintkeznek a szervezettől idegen antigénekkel. Az ilyen érintkezés elsősorban a limfoid szövetben, az antigén belépési helyén történik. Az érintkezés után a klónok elszaporodnak(a görög klon szóból - csíra, utódok)Az erre az antigénre specifikus T- és B-sejtek, valamint e klónok sejtjeinek többségének differenciálódása rövid életű végső effektorokká (T-effektorok a T-limfocitákból és plazmasejtek a B-limfocitákból). Ezen antigén-specifikus klónok T- és B-limfocitái közül néhány anélkül szaporodik, hogy rövid életű effektor klónokká válna. immunológiai memóriasejtek. Ez utóbbiak részben más másodlagos limfoid szervekbe vándorolnak, aminek következtében megnövekedett limfociták jelennek meg bennük, amelyek arra az antigénre specifikusak, amelyet legalább egyszer megtámadtak a szervezetben. Ez immunológiai memóriát hoz létre egy adott antigén számára az immunrendszer egészében.
A limfociták áramlását a véráramból a másodlagos limfoid szervekbe szigorúan szabályozzák. Az érett T- és B-limfociták jelentős részeegyértelműen kering a véráramban a nyirokszervek között (ún recirkuláló limfociták). A limfociták újrahasznosítása a limfociták vérből az immunrendszer szerveibe, a perifériás szövetekbe és vissza a vérbe vándorlásának folyamatát jelenti (8.4. ábra). A limfocitáknak csak egy kis része tartozik a nem recirkuláló készletbe.
A limfociták újrahasznosításának funkcionális célja a testszövetek folyamatos „immunfelügyelete” immunkompetens limfociták által, az idegen és megváltozott saját antigének hatékony kimutatása, valamint a limfocitopoézis szerveinek tájékoztatása az antigének különböző szövetekben való megjelenéséről. Létezik gyors (néhány órán belül végrehajtott) recirkuláció és lassú recirkuláció (hetekig tart). A gyors recirkuláció során a vér limfociták specifikusan a nyirokszervekben elhelyezkedő speciális erek falához kötődnek - magas endotéliummal rendelkező posztkapilláris venulák -, majd ezeken az endothelsejteken keresztül a limfoid szövetbe, majd a nyirokerekbe és a mellkasi nyirokereken keresztül vándorolnak. visszavezetni a vérbe. A mellkasi vezetékben található limfociták körülbelül 90%-a vándorol ezen az úton. A lassú recirkuláció során a vér limfociták a nem immunszervekre jellemző, laphám endotéliummal ellátott posztkapilláris venulákon keresztül különböző perifériás szövetekbe vándorolnak, majd a nyirokerekbe, nyirokcsomókba, majd a nyirokáramláson keresztül a mellkasi nyirokcsatornába ismét a vérbe jutnak. . A mellkasi vezetékben található limfociták hozzávetőleg 5-10%-a kerül újrahasznosításra ezen a módon.
A limfociták specifikus kötődése a magas endotéliummal rendelkező posztkapilláris venulák falához annak köszönhető, hogy az endothelsejtek felszínén bizonyos molekulák és a megfelelő receptorok a T- és B-limfocitákon jelen vannak (8.5. ábra). Ez a mechanizmus biztosítja a limfociták bizonyos populációinak szelektív felhalmozódását a nyirokcsomókban és más másodlagos limfoid szervekben. A Peyer tapaszok körülbelül 70% B-limfocitát és 10-20% T-limfocitát tartalmaznak, míg a perifériás nyirokcsomókban körülbelül 70% T- és 20% B-sejteket. Számos antigén által aktivált T- és B-limfocita elhagyja azt a helyet, ahol aktiválódott, majd a véráramban keringve visszatér ugyanabba a vagy hasonló limfoid szervbe. Ez a minta az alapja helyi immunitás szervek és szövetek. A recirkuláló limfociták közül a legtöbb
Mindkét típusú T-limfociták és immunológiai memóriasejtek migrációs sebességgel rendelkeznek.
A bőr és a nyálkahártya sejtjei is közvetlenül részt vesznek az immunvédelemben, mechanikai akadályt képezve az idegen antigénnel szemben. Mint mechanikai tényezők nem specifikus védekező mechanizmusok szóba jöhet a többrétegű hám felszíni rétegeinek sejtjeinek hámlása (hámlása), a nyálkahártyát borító nyálkaképződés, a hám felszínén a nyálkát szállító csillók verése (a légutakban - mukociliáris transzport) . A mikrobákat a nyál, a könny, a vizelet és más folyadékok áramlása is eltávolítja a hám felszínéről.
NAK NEK humorális immunkomponensek immunológiailag aktív molekulák széles skáláját foglalják magukban, az egyszerűtől a nagyon összetettig, amelyeket immunkompetens és más sejtek termelnek, és részt vesznek a szervezet idegen vagy hibás anyagokkal szembeni védelmében. Közülük mindenekelőtt a fehérje jellegű anyagokat kell kiemelni - immunglobulinok, citokinek, komplement komponensek rendszere, akut fázisú fehérjék, interferon és mások. Az immunkomponensek közé tartoznak az enzimgátlók, amelyek elnyomják a baktériumok enzimaktivitását, a vírusgátlók és számos kis molekulájú anyag, amelyek az immunreakciók mediátorai (hisztamin, szerotonin, prosztaglandinok és mások). A szervezet hatékony védelmében nagy jelentőséggel bír a szövetek oxigénnel való telítettsége, a környezet pH-ja, a Ca 2+ jelenléte, ill. Mg 2+ és egyéb ionok, nyomelemek, vitaminok stb.
8. 2. A NEM SPECIFIKUS (VELÉLETETETT) IMUNITÁS MECHANIZMUSAI
Nem specifikus (veleszületett) védekező mechanizmusok Mindazon élettani tényezők kombinációját jelentik, amelyek a) megakadályozhatják a szervezetbe jutást, vagy b) semlegesíthetik és elpusztíthatják a behatolt idegen anyagokat, részecskéket vagy a benne képződött saját módosult sejteket. Ezek a mechanizmusok nem specifikusak a ható szerre.
Az említett mechanikai és kémiai tényezőkön kívül számos egyéb védekezési mód is létezik: fagocitózis(sejtek általi „evés”), a vírussal fertőzött és daganatos sejtek extracelluláris elpusztítása citotoxikus faktorok segítségével (celluláris citotoxicitás)és az idegen sejtek elpusztítása oldható baktericid vegyületek segítségével.
Az immunrendszer különféle összetevőkből áll - szervekből, szövetekből és sejtekből, amelyek a funkcionális kritérium (a szervezet immunvédelmének megvalósítása) és a szervezet anatómiai és fiziológiai elve (szerv-keringési elv) szerint ebbe a rendszerbe sorolhatók. Az immunrendszer megkülönbözteti az elsődleges szerveket (csontvelő és csecsemőmirigy), a másodlagos szerveket (lép, nyirokcsomók, Peyer-foltok stb.), Valamint a diffúz elhelyezkedésű nyirokszöveteket - az egyes limfoid tüszőket és azok klasztereit. Különösen megkülönböztethető a nyálkahártyákkal kapcsolatos limfoid szövet (Nyálkahártyával kapcsolatos nyirokcsomó - MALÁTA).
Limfoid rendszer- limfoid sejtek és szervek gyűjteménye. A limfoid rendszert gyakran az immunrendszer anatómiai megfelelőjeként és szinonimájaként emlegetik, de ez nem teljesen igaz. A limfoid rendszer csak egy része az immunrendszernek: az immunrendszer sejtjei a nyirokereken keresztül a nyirokszervekbe vándorolnak - az immunválasz indukciójának és kialakulásának helyére. Ezenkívül a nyirokrendszert nem szabad összetéveszteni a nyirokrendszerrel - a nyirokrendszerrel, amelyen keresztül a nyirok kering a szervezetben. A nyirokrendszer szorosan kapcsolódik a keringési és endokrin rendszerhez, valamint az integumentáris szövetekhez - a nyálkahártyákhoz és a bőrhöz. A nevezett rendszerek azok a fő partnerek, amelyekre az immunrendszer támaszkodik munkájában.
Az immunrendszer szerveződésének szervi-keringési elve. Egy felnőtt egészséges ember szervezete körülbelül 10 13 limfocitát tartalmaz, azaz. a testben körülbelül minden tizedik sejt limfocita. Anatómiailag és élettanilag az immunrendszer szerv-keringési elv szerint szerveződik. Ez azt jelenti, hogy a limfociták nem szigorúan rezidens sejtek, hanem intenzíven keringenek a limfoid szervek és a nem limfoid szövetek között a nyirokereken és a véren keresztül. Így ≈10 9 limfocita halad át minden nyirokcsomón 1 óra alatt. A limfociták vándorlását az határozza meg
specifikus molekulák specifikus kölcsönhatásai a limfociták membránjain és az érfal endothelsejtjein [az ilyen molekulákat adhezineknek, szelektineknek, integrinnek, homing receptoroknak nevezik (az angolból. itthon- otthon, a limfocita lakóhelye)]. Ennek eredményeként minden szerv rendelkezik egy jellegzetes limfocitapopuláció-készlettel és immunválasz-partnersejtjeivel.
Az immunrendszer összetétele. A szervezet típusától függően az immunrendszer különböző szerveit és szöveteit különböztetjük meg (2-1. ábra).
. Hematopoetikus csontvelő - hematopoietikus őssejtek (HSC) lokalizációjának helye.
Rizs. 2-1. Az immunrendszer összetevői
. Kapszulázott szervek: csecsemőmirigy, lép, nyirokcsomók.
. Kapszulázatlan limfoid szövet.
-Nyálkahártyák limfoid szövetei(MALT - Nyálkahártyával összefüggő limfoid szövet). A helytől függetlenül a nyálkahártya intraepiteliális limfocitáit, valamint speciális képződményeket tartalmaz:
◊ az emésztőrendszerhez kapcsolódó limfoid szövet (GALT - Bélhez kapcsolódó limfoid szövet). Tartalmaz mandulákat, vakbelet, Peyer-foltokat, lamina propria("lamina propria"), az egyes limfoid tüszők és csoportjaik;
◊ hörgőkhöz és hörgőcsövekhez kapcsolódó limfoid szövet (BALT - Bronchus-asszociált limfoid szövet);
◊lymphoid szövet, amely a női nemi szervekhez kapcsolódik (VALT - Vulvovaginális eredetű limfoid szövet);
◊orrgarathoz kapcsolódó limfoid szövet (NALT - Az orrhoz kapcsolódó limfoid szövet e).
A máj különleges helyet foglal el az immunrendszerben. Limfociták és az immunrendszer egyéb sejtjei olyan alpopulációkat tartalmaznak, amelyek limfoid gátként a portális véna vérét „szolgálják”, amely a bélben felszívódó összes anyagot hordozza.
A bőr limfoid alrendszere - bőrrel összefüggő limfoid szövet (SÓ - Bőrrel összefüggő limfoid szövet)- disszeminált intraepiteliális limfociták és regionális nyirokcsomók és nyirokelvezető erek.
. Perifériás vér - az immunrendszer szállítási és kommunikációs összetevője.
Az immunrendszer központi és perifériás szervei
. Központi hatóságok. A vérképző csontvelő és a csecsemőmirigy az immunrendszer központi szervei, ahol a myelopoiesis és lymphopoiesis kezdődik – a monociták és limfociták differenciálódása HSC-ből érett sejtekké.
A magzat születése előtt a magzati májban B-limfociták fejlődnek ki. Születés után ez a funkció átkerül a csontvelőbe.
A csontvelőben az erythropoiesis (vörösvérsejtek képződése), a myelopoiesis (neutrofilek képződése,
monociták, eozinofilek, bazofilek), megakariocitopoiesis (thrombocytaképződés), valamint a DC-k, NK-sejtek és B-limfociták differenciálódása is végbemegy. - A T-limfociták prekurzorai a csontvelőből a csecsemőmirigybe és az emésztőrendszer nyálkahártyájába vándorolnak, hogy limfopoézisen (extrathymiás fejlődésen) menjenek keresztül.
. Perifériás szervek. A perifériás limfoid szervekben (lép, nyirokcsomók, nem kapszulázott limfoid szövet) az érett naiv limfociták érintkezésbe kerülnek antigénnel és APC-vel. Ha egy limfocita antigénfelismerő receptora egy perifériás limfoid szervben komplementer antigént köt, akkor a limfocita immunválasz módban a további differenciálódás útjára lép, azaz. szaporodni kezd és effektormolekulákat termelnek - citokinek, perforin, granzimek stb. A limfociták ezen további differenciálódását a periférián ún. immunogenezis. Az immunogenezis eredményeként effektor limfociták klónjai képződnek, amelyek felismerik az antigént, és megszervezik mind saját, mind a szervezet perifériás szöveteinek pusztítását, ahol ez az antigén jelen van.
Az immunrendszer sejtjei. Az immunrendszer különböző eredetű - mezenchimális, ekto- és endodermális - sejteket foglal magában.
. Mesenchymalis eredetű sejtek. Ide tartoznak a limfo/hematopoiesis prekurzoraitól differenciált sejtek. Fajták limfociták- T, B és NK, amelyek az immunválasz során együttműködnek különböző leukociták - monociták/makrofágok, neutrofilek, eozinofilek, bazofilek, valamint DC-k, hízósejtek és vaszkuláris endoteliális sejtek. Még vörös vérsejtek hozzájárulnak az immunválasz megvalósításához: „antigén-antitest-komplement” immunkomplexeket szállítanak a májba és a lépbe fagocitózis és pusztulás céljából.
. Hámszövet. Egyes limfoid szervek (csecsemőmirigy, néhány nem kapszulázott limfoid szövet) ektodermális és endodermális eredetű hámsejteket tartalmaznak.
Humorális tényezők. A sejtek mellett az „immunanyagot” oldható molekulák - humorális tényezők - képviselik. Ezek a B-limfociták termékei - antitestek (más néven immunglobulinok) és az intercelluláris kölcsönhatások oldható mediátorai - citokinek.
THIMUSZ
A csecsemőmirigyben (csecsemőmirigy) A T-limfociták jelentős részének limfopoézise megy végbe (a „T” a szóból származik Thymus). A csecsemőmirigy 2 lebenyből áll, mindegyiket kötőszövet kapszula veszi körül. A kapszulából kinyúló válaszfalak lebenyekre osztják a csecsemőmirigyet. A csecsemőmirigy minden lebenyében (2-2. ábra) 2 zóna van: a periféria mentén - corticalis (cortex), a központban - agyi (csontvelő). A szerv térfogatát epiteliális keret tölti ki (hámszövet), amelyben elhelyezkednek timociták(a csecsemőmirigy éretlen T-limfocitái), DKÉs makrofágok. A DC-k túlnyomórészt a kortikális és az agyi régiók közötti átmeneti zónában helyezkednek el. A makrofágok minden zónában jelen vannak.
. Hámsejtek folyamataik körülveszik a csecsemőmirigy-limfocitákat (timocitákat), ezért nevezik őket "ápolósejtek"("ápoló" cellák vagy "dada" cellák). Ezek a sejtek nemcsak támogatják a timociták fejlődését, hanem termelnek is

Rizs. 2-2. A csecsemőmirigy lebenyének felépítése
citokinek IL-1, IL-3, IL-6, IL-7, LIF, GM-CSF és expressz adhéziós molekulák LFA-3 és ICAM-1, amelyek komplementerek a timociták felületén lévő adhéziós molekulákkal (CD2 és LFA-1) . A lebenyek medulláris zónájában csavart hámsejtek sűrű képződményei vannak - Hassall holttestét(csecsemőmirigytestek) - a degeneráló hámsejtek kompakt felhalmozódásának helyei.
. Timociták megkülönböztetni a csontvelői SCC-ktől. A timocitákból a differenciálódási folyamat során T-limfociták képződnek, amelyek MHC-vel kombinálva képesek felismerni az antigéneket. A legtöbb T-limfocita azonban vagy nem rendelkezik ezzel a tulajdonsággal, vagy felismeri a saját antigéneket. Az ilyen sejtek perifériára való felszabadulásának megakadályozására a csecsemőmirigyben az apoptózis indukálásával megindul az eliminációjuk. Normális esetben tehát a csecsemőmirigyből csak azok a sejtek kerülnek a keringésbe, amelyek képesek felismerni az antigéneket „a maguk” MHC-jával kombinálva, de nem váltanak ki autoimmun reakciókat.
. Hematotímiás gát. A csecsemőmirigy erősen vaszkularizált. A kapillárisok és venulák falai hematotímiás gátat képeznek a csecsemőmirigy bejáratánál, és esetleg a kilépésnél. Az érett limfociták vagy szabadon lépnek ki a csecsemőmirigyből, mivel minden lebenyben van egy efferens nyirokér, amely a nyirokot a mediastinum nyirokcsomóihoz szállítja, vagy a kortikomedulláris régióban magas endotéliummal rendelkező posztkapilláris venulák falán keresztül és/vagy a nyirokcsomók falán keresztül. közönséges vérkapillárisok.
. Az életkorral összefüggő változások. A születés idejére a csecsemőmirigy teljesen kialakul. A timociták sűrűn benépesítik egész gyermekkorban és egészen a pubertásig. A pubertás után a csecsemőmirigy mérete csökkenni kezd. A thymectomia felnőtteknél nem vezet súlyos immunrendszeri zavarokhoz, mivel gyermek- és serdülőkorban a perifériás T-limfociták szükséges és elegendő készlete jön létre az élet hátralévő részében.
A NYIROKCSOMÓK
A nyirokcsomók (2-3. ábra) többszörös, szimmetrikusan elhelyezkedő, kapszulázott perifériás nyirokszervek, bab alakúak, mérete 0,5-1,5 cm hosszúságú (gyulladás hiányában). A nyirokcsomók afferens (afferens) nyirokereken keresztül szívják el a szövetet (minden csomóhoz több van).

Rizs. 2-3. Az egér nyirokcsomójának szerkezete: a - corticalis és medulla részek. A kérgi részen nyiroktüszők találhatók, amelyekből agyzsinórok nyúlnak be a medullába; b - a T- és B-limfociták eloszlása. A csecsemőmirigy-függő zóna rózsaszínnel, a csecsemőmirigy-független zóna sárgával van kiemelve. A T-limfociták a posztkapilláris venulákból lépnek be a csomópont parenchymájába, és érintkezésbe kerülnek follikuláris dendritikus sejtekkel és B-limfocitákkal
új folyadék. Így a nyirokcsomók „szokások” minden anyag számára, beleértve az antigéneket is. Az egyetlen efferens (kiáramló) ér a csomópont anatómiai kapujából jön ki, az artériával és a vénával együtt. Ennek eredményeként a nyirok bejut a mellkasi nyirokcsatornába. A nyirokcsomó parenchimája T-sejtből, B-sejtes zónákból és medulláris zsinórokból áll.
. B-sejtes zóna. A kéreg kötőszöveti trabekulák által sugárirányú szektorokra van osztva, és limfoid tüszőket tartalmaz, ez a B-limfocita zóna. A tüszők stromája follikuláris dendritikus sejteket (FDC) tartalmaz, amelyek egy speciális mikrokörnyezetet alkotnak, amelyben egyedülálló folyamat megy végbe a B-limfociták számára, az immunglobulin gének variábilis szegmenseinek szomatikus hipermutagenezise és az antitestek leginkább affinitású változatainak szelekciója ("antitest affinitás-érés" ”). A limfoid tüszők 3 fejlődési szakaszon mennek keresztül. Elsődleges tüsző- naiv B-limfocitákat tartalmazó kis tüsző. Miután a B limfociták belépnek az immunogenezisbe, a csíra (csíra) központ, intenzíven szaporodó B-sejteket tartalmaz (ez körülbelül 4-5 nappal az aktív immunizálás után következik be). Ez másodlagos tüsző. Az immunogenezis befejeztével a limfoid tüsző mérete jelentősen csökken.
. T sejt zóna. A nyirokcsomó paracorticalis (T-dependens) zónájában csontvelő eredetű T-limfociták és interdigitális DC-k találhatók (ezek különböznek az FDC-ktől), amelyek antigéneket mutatnak be a T-limfocitáknak. A magas endotéliummal rendelkező posztkapilláris venulák falán keresztül a limfociták a vérből a nyirokcsomókba vándorolnak.
. Agyzsinórok. A parakortikális zóna alatt makrofágokat tartalmazó medulláris zsinórok találhatók. Aktív immunválasz mellett sok érett B-limfocita - plazmasejt - látható ezekben a szálakban. A zsinórok a medulla sinusába folynak, ahonnan az efferens nyirokerek lépnek ki.
LÉP
Lép- egy viszonylag nagy párosítatlan szerv, amelynek tömege körülbelül 150 g, a lép nyirokszövete. fehér pép. A lép a vérbe jutó antigének limfocita „vámháza”. Limfociták

Rizs. 2-4. Emberi lép. A lép csecsemőmirigy-függő és csecsemőmirigy-független zónái. A T-limfociták (zöldsejtek) felhalmozódása a trabekulákból kilépő artériák körül csecsemőmirigy-függő zónát alkot. A nyiroktüsző és a környező fehér pulpa limfoid szövet csecsemőmirigy-független zónát alkot. A nyirokcsomók tüszőihez hasonlóan vannak B-limfociták (sárga sejtek) és follikuláris dendrites sejtek. A másodlagos tüsző egy csíraközpontot tartalmaz gyorsan osztódó B-limfocitákkal, amelyeket kis nyugalmi limfociták gyűrűje vesz körül (köpeny).
lépek gyűlnek össze az arteriolák körül úgynevezett periarterioláris csatolások formájában (2-4. ábra).
A T kapuzatú csatolási zóna közvetlenül körülveszi az arteriolát. A B-sejtes tüszők közelebb helyezkednek el a muff széléhez. A lép arteriolái a szinuszoidokba áramlanak (ez már vörös pép). A sinusoidok venulákban végződnek, amelyek a lépvénában gyűlnek össze, amely a vért a máj portális vénájába szállítja. A vörös és fehér pépet egy diffúz marginális zóna választja el, amelyet B-limfociták (marginális B-sejtek) és speciális makrofágok speciális populációja népesít be. A marginális zóna sejtek fontos kapcsot jelentenek a veleszületett és szerzett immunitás között. Itt történik a szervezett limfoid szövet legelső érintkezése a vérben keringő lehetséges kórokozókkal.
MÁJ
A máj fontos immunfunkciókat lát el, ami a következő tényekből következik:
A máj a limfopoézis erőteljes szerve az embrionális időszakban;
Az allogén májátültetések kevésbé gyorsan kilökődnek, mint más szervek;
Az orálisan beadott antigénekkel szembeni tolerancia csak a máj normál fiziológiás vérellátása mellett indukálható, és nem váltható ki műtét után portocaval anasztomózisok létrehozására;
A máj akut fázisú fehérjéket (CRP, MBL stb.), valamint a komplementrendszer fehérjéit szintetizálja;
A máj különböző limfociták alpopulációkat tartalmaz, köztük egyedi limfocitákat, amelyek egyesítik a T- és NK-sejtek (NKT-sejtek) jellemzőit.
A máj sejtes összetétele
Hepatociták a máj parenchimáját alkotják, és nagyon kevés MHC-I molekulát tartalmaznak. A hepatociták általában szinte egyáltalán nem hordoznak MHC-II molekulát, de expressziójuk fokozódhat májbetegségekben.
Kupffer sejtek - máj makrofágok. A májsejtek teljes számának körülbelül 15%-át és a szervezet összes makrofágjának 80%-át teszik ki. A makrofágok sűrűsége nagyobb a periportális területeken.
Endothel A máj szinuszoidjainak nincs alapmembránja - egy vékony extracelluláris szerkezet, amely különböző típusú kollagénekből és más fehérjékből áll. Az endothelsejtek lumenekkel rendelkező monoréteget alkotnak, amelyen keresztül a limfociták közvetlenül érintkezhetnek a májsejtekkel. Ezenkívül az endothel sejtek különféle scavenger receptorokat expresszálnak. (scavenger receptorok).
Limfoid rendszer A máj a limfocitákon kívül a nyirokkeringés egy anatómiai szakaszát is tartalmazza - a Disse terét. Ezek a terek egyrészt közvetlenül érintkeznek a máj sinusoidjainak vérével, másrészt a májsejtekkel. A májban a nyirokáramlás jelentős – a szervezet teljes nyirokáramlásának legalább 15-20%-a.
Csillagsejtek (Ito-sejtek) a Disse terekben található. Zsírvakuolákat tartalmaznak A-vitaminnal, valamint a simaizomsejtekre jellemző α-aktint és dezmint. A csillagsejtek miofibroblasztokká alakulhatnak át.
A NYÁK MEMBRÁNOK ÉS A BŐR NYÁKSZÖVETE
A nyálkahártya nem kapszulázott limfoid szövetét a Pirogov-Waldeyer garat limfoid gyűrűje, a vékonybél Peyer-foltjai, a vakbél limfoid tüszői, a gyomor, a belek, a hörgők és a hörgők nyálkahártyájának limfoid szövetei képviselik, az urogenitális rendszer szervei és más nyálkahártyák.
Peyer foltjai(2-5. ábra) - ben elhelyezkedő csoportos nyiroktüszők lamina propria vékonybél. A tüszők, pontosabban a tüszők T-sejtjei a bélhám mellett helyezkednek el az úgynevezett M-sejtek alatt („M” membrán, ezekben a sejtekben nincsenek mikrobolyhok), amelyek a Peyer-tapasz „bejárati kapuját” jelentik. A limfociták nagy része csíraközpontokkal rendelkező B-sejtes tüszőkben található. T-sejtes zónák veszik körül a tüszőt a hámhoz közelebb. A B-limfociták 50-70%-át teszik ki, a T-limfociták - 10-30%-át az összes Peyer-tapasz sejtnek. A Peyer tapaszok fő funkciója a B-limfociták immunogenezisének és differenciálódásának fenntartása.

Rizs. 2-5. Peyer-folt a bélfalban: a - általános kép; b - egyszerűsített diagram; 1 - enterociták (bélhám); 2 - M-sejtek; 3 - T-sejt zóna; 4 - B-sejt zóna; 5 - tüsző. A szerkezetek közötti léptéket nem tartják be
antitesteket - túlnyomórészt szekréciós IgA-t - termelő plazmasejtekbe vándorolnak. Az IgA termelése a bélnyálkahártyában a szervezet teljes napi immunglobulin-termelésének több mint 70%-át teszi ki – felnőtteknél naponta körülbelül 3 g IgA. A szervezet által szintetizált összes IgA több mint 90%-a a nyálkahártyán keresztül kiválasztódik a bél lumenébe.
Intraepiteliális limfociták. A nyálkahártyák a szervezett limfoid szöveten kívül egyetlen intraepiteliális T-limfocitát is tartalmaznak, amelyek a hámsejtek között szétszóródnak. Felületükön egy speciális molekula expresszálódik, amely biztosítja ezeknek a limfocitáknak az enterocitákhoz való tapadását - az α E (CD103) integrin. Az intraepiteliális limfociták körülbelül 10-50%-a TCRγδ + CD8αα + T limfociták.



