A citológiában alkalmazott kutatási módszerek. Módszerek a sejtek tanulmányozására. Módszerek élő sejtek és szövetek vizsgálatára
A sejtek tanulmányozására szolgáló morfo-funkcionális módszer fő jellemzője az a vágy, hogy megértsük az adott funkciót meghatározó biokémiai folyamatok szerkezeti alapjait, vagyis ezeket a folyamatokat specifikus sejtstruktúrákkal összekapcsoljuk.
Ezzel a módszerrel a végső cél megegyezik a molekuláris biológia és a sejtszerkezeti biokémia által követett céllal. Az e tudományok által egy közös probléma megoldására alkalmazott módszerek azonban alapvetően különböznek egymástól. Ha a molekuláris biológiában és a szerkezeti biokémiában nélkülözhetetlen feltétel a sejt elpusztítása és a vizsgált szerkezet többé-kevésbé tiszta frakció formájában történő izolálása, akkor a citológiai vizsgálatokban éppen ellenkezőleg, a sejt megőrzése az előfeltétel. a sejt integritása. Ebben az esetben törekedni kell a külső interferencia minimálisra csökkentésére, és meg kell próbálni tanulmányozni egyes komponensek szerkezeti és biokémiai szerveződését a teljes sejtrendszeren belül.
A morfofunkcionális irány kutatása gyorsan fejlődött az elmúlt évtizedekben. Ebben az időben számos alapvetően új módszert fejlesztettek ki a sejtszerkezetek kvalitatív és kvantitatív elemzésére. Ez a megközelítés szorosan kapcsolódik a biológiai tudományok új ágaihoz, és különösen a molekuláris biológiához, ami meghatározza, hogy az ilyen kutatások igen jelentős mértékben járulnak hozzá a sejtszerveződés általános törvényszerűségeire vonatkozó ismereteink fejlődéséhez.
Elektronmikroszkópia
Az egyik legelterjedtebb, a szerkezeti és biokémiai vizsgálatokban már klasszikussá vált módszer az elektronmikroszkópos módszer annak különféle módosításaiban. Ezek a módosítások egyrészt a vizsgált struktúrák elemzésének eltérő megközelítéséből, másrészt a sejtek ultrastrukturális vizsgálatokra való felkészítésének sajátosságaiból fakadnak. A hagyományos transzmissziós (transzmissziós) mikroszkópok nagy felbontása lehetővé teszi nemcsak a nukleáris és citoplazmatikus apparátus összes organellumának elemzését, hanem egyes szupramolekuláris szerveződési szinten elhelyezkedő struktúrák elemzését is, például hordozó és kontraktilis mikrofibrillumok, mikrotubulusok és néhány multienzim komplexek. Jelenleg a nagyfeszültségű elektronmikroszkópos módszert egyre sikeresebben alkalmazzák a sejtek szerveződésének szisztémás és alrendszeri szintjén történő tanulmányozására. A transzmissziós elektronmikroszkóphoz képest az áthatoló elektronsugár jóval nagyobb energiája miatt ez a módszer lehetővé teszi „vastag” metszetek, vagy akár egész szétterített sejtek mikroszkóp alatti tanulmányozását, ami lehetővé teszi például az átfogó elemzését. a sejtfelszíni apparátus szubmembrán rostjainak komplex rendszere.
A sejt felszíni apparátusának működésének, a sejtmag felszíni apparátusának egyes alrendszereinek kapcsolatának és számos más általános citológiai kérdésnek, a pásztázó elektronmikroszkópos módszernek a tanulmányozásában, amely lehetővé teszi a egy tárgy felülete térfogatban jelentős jelentőséget kap.
Freeze-chip módszer
A morfobiokémiai irány citológiai vizsgálatában különleges és alapvetően fontos helyet foglal el a fagyasztás-hasítás módszer. Ez a legkíméletesebb módszer a biológiai objektumok ultrastrukturális elemzésre való előkészítésére, azaz minimális változást okoz a sejtszerkezetekben a natív állapotukhoz képest. A módszer lényege a következő. A tárgyat folyékony nitrogén atmoszférába helyezik, amely azonnal leállítja az összes anyagcsere-folyamatot. Ezután a fagyott tárgyból chips készül. A replikákat a forgácsok felületéről úgy nyerik, hogy fémfóliát helyeznek rájuk. Ezeket a filmeket ezt követően elektronmikroszkóp alatt vizsgálják. A fagyasztási-hasítási módszer előnye, hogy a hasítási sík általában áthalad a membrán hidrofób fázisán, és ez lehetővé teszi az integrált membránfehérjék mennyiségének, méretének és elrendeződésének, azaz közvetlenül a belső morfobiokémiai szerveződésnek a tanulmányozását. membránok, a hasításokon. A módszer igen értékes eredményeket adott különféle típusú membránszerkezetek és speciális képződmények, például bizonyos típusú sejtkontaktusok vizsgálatában.
Citokémiai módszer
A citológiai kutatás szerkezeti-biokémiai aspektusának fő feladatában - a struktúrák funkcionális jelentőségének feltárásában biokémiai szerveződésük elemzésén keresztül - a citokémiai módszerek rendkívül fontos szerepet töltenek be. Jelenleg folyamatosan fejlesztik ezeket mind a vizsgált szerkezetekben található kémiai vegyületek pontos minőségi azonosítása, mind pedig mennyiségi értékelésük szempontjából. A kvantitatív citospektrofotometriát lehetővé tevő speciális műszerek segítségével nem csak a sejt egészében, hanem a sejtmag vagy citoplazmatikus struktúrák szintjén is meg lehet határozni egy adott anyag, például RNS és DNS tartalmát. Az interferencia-mikroszkópiának köszönhetően felmérhető a sejtben lévő fehérje teljes mennyisége és annak élete során bekövetkezett változásai.
Létezik egy módszer az enzimek citokémiai azonosítására, amely lehetővé teszi nemcsak egy adott vegyület lokalizációjának és mennyiségének megítélését a sejtszerkezetekben, hanem ezen vegyületek szintézisének és intracelluláris transzportjának folyamatait is.
Az enzimek citokémiája a szubsztrát-enzim kölcsönhatás elvén alapul, marker vegyületek segítségével, amelyek kicsapódnak. Az enzimrendszerek lokalizációjának, esetenként aktivitásának meghatározásával meg tudjuk ítélni bizonyos biokémiai folyamatok lokalizációját a sejtszerkezetekben.
Autoradiográfia
Az autoradiográfiás módszer az enzimek citokémiájához hasonlóan lehetőséget ad az intracelluláris szintézis és transzport vizsgálatára, ugyanakkor még szélesebb lehetőségeket rejt magában. Az autográf módszer a radioaktív prekurzorok felhasználásán alapul mesterséges izotópokkal (3 H, 14 C, 35 S stb.) jelölt makromolekulák szintézisére. Nemcsak bizonyos makromolekulák szintézisének helyeinek lokalizálását teszi lehetővé, hanem ezen vegyületek sejten belüli transzportjának specifikus útvonalainak nyomon követését is, hogy relatív kvantitatív értékelést adjon a szintézis intenzitásáról és a makromolekulák mozgási sebességéről a sejtszerkezetekben. Ily módon különösen először mutatták meg az RNS mozgását a sejtmagból a sejtek citoplazmájába, részletesen nyomon követték a szekréciós sejtekben a szintézis lokalizációját és a váladék intracelluláris transzportját, és sok más, általánosan fontos tényt. citológiát tártak fel. Lényegében ez a módszer az egyik legtipikusabb, a strukturális-biokémiai kutatási irányra jellemző módszer, hiszen lehetővé teszi az intracelluláris struktúrákban zajló anyagcsere-folyamatok közvetlen vizsgálatát egy ép, sértetlen (a biokémiai vizsgálatokhoz hasonlóan) sejtben. A módszer lényege a mesterséges izotóppal jelölt molekulák fényképészeti emulzió segítségével történő kimutatásán alapul, amely a jelölt prekurzor bevezetése után különböző időpontokban rögzített sejt- és szövetmetszeteket fed le.
Immuncitokémiai módszer
Jelenleg a sejtszerkezetek egyes fehérjéinek nagyon pontos kvalitatív elemzése lehetséges a teljes sejtrendszeren belül. Ezt az elemzést immuncitokémiai módszerekkel végezzük. Ezeknek a módszereknek a lényege, hogy egy specifikus fehérje antigénként szolgál, amelyre bármely emlős szervezetében specifikus antitestek képződnek. Ez utóbbiakat fluoreszcens festékkel vagy más jelzőanyaggal kombinálják. Ezután a vizsgált sejtet jelölt antitesteket tartalmazó szérummal kezelik. A specifikus jelölt antitestek szigorúan szelektíven kötődnek a vizsgált fehérjéket tartalmazó struktúrákhoz. Ezzel a módszerrel különösen az aktin-miozin rendszer fő és kiegészítő kontraktilis fehérjéinek lokalizációját tárták fel a sejtek submembrán fibrilláris apparátusában, és eloszlásuk módosulását a mitotikus apparátus kialakulása és a citotomia folyamata során. mutatták be. Ugyanezt a módszert sikeresen alkalmaztuk a membrán szerveződés folyadék-mozaik modelljének érvényességének bizonyítására.
Átfogó sejtkutatási módszerek
Az utóbbi időben különösen nagy sikereket értek el a sejtek szerkezeti és biokémiai szerveződésének vizsgálatában az ultrastrukturális elemzési módszerek, valamint a citokémiai és autoradiográfiai módszerek integrált alkalmazásával. Ezek a sikerek elsősorban az ultrastrukturális szintű citokémia és autoradiográfia speciális módszereinek kifejlesztésének köszönhetőek, amelyek lehetővé teszik az anyagcsere folyamatok közvetlen elemzését a sejtszerveződés megnevezett szintjén, a biokémiai folyamatok „strukturálását”, a sejtek sajátos jelentésének megismerését. bizonyos sejtszerkezetek az intracelluláris anyagcsere összetett folyamatainak egyes kapcsolataiban. Ebben a tekintetben kiterjedt anyag halmozódott fel a citoplazma membránfázisának különböző változatainak szerepéről a szintetikus anabolikus folyamatokban és az intracelluláris katabolizmus folyamataiban.
Különösen a sejtek lizoszómális apparátusának szerveződésének és működésének tanulmányozása terén értek el jelentős sikereket. A sejtek nukleáris apparátusának vizsgálata során fontos új tényekre jutottak. Citokémiai módszerekkel ultrastrukturális szinten lehet azonosítani a ribonukleoproteineket (RNP-k) és dezoxiribonukleoproteineket (DNP-ket), és ezzel jelentős előrelépést lehet elérni a különböző típusú RNP-k transzkripciójának, érésének és intranukleáris transzportjának eukarióta sejtekben történő megszervezésének, valamint felhasználásának tanulmányozásában. Az elektronikus autoradiográfia lehetővé tette az egyes sejtstruktúrák szerepének részletezését ezekben a folyamatokban. Például részletesen tanulmányozható volt a nucleolus működése, és specifikusan strukturálható a riboszómális RNS képződési folyamata benne.
A molekuláris biológiai és szerkezeti biokémiai szempontok és módszerek ilyen szintézise nagyon jellemző az egyes sejtkomponensek finom szerveződésével kapcsolatos számos más fontos kérdés kidolgozására is. Ugyanakkor a molekuláris biológiai és morfobiokémiai citológiai analízis szoros kapcsolata nemcsak a végeredmények szintézisében, hanem magában a kutatási folyamat során való kölcsönhatásban is megnyilvánul. Az ilyen interakciót vagy biokémiai és citológiai módszerekkel végzett komplex munka elvégzésével, speciális biokémikusok és citológusok által, vagy speciális komplex módszerek alkalmazásával, amelyek a sejtszerkezetek biokémiai és citológiai elemzésének határán vannak.
Az első típusra példa a sejtkomponensek biokémiai izolálási módszereinek és azok finom ultrastrukturális elemzésének kombinációja. Ily módon először készültek fényképek a működő génekről a DNS, az RNS polimerázok és az átírt RNS-molekulák azonosításával. Ennek a módszernek a továbbfejlesztése bizonyos esetekben lehetővé teszi a transzkripció intenzitásának figyelembevételét az RNS-polimeráz komplexek számának közvetlen megszámlálásával. Elektronmikroszkóp segítségével közvetlenül tanulmányozhatja a DNS-replikáció mintázatait biokémiai módszerekkel izolált cirkuláris vagy lineáris DNS-molekulákon. Az ultrastrukturális elemzési módszereket széles körben alkalmazzák az egyes fehérjék lokalizációjának immuncitokémiai vizsgálataiban, riboszomális szubpartikulumokban, a DNP különböző szintű szerveződésének vizsgálatában és sok más esetben is.
A speciálisan kifejlesztett komplex módszerek tipikus példája a DNS és RNS hibridizációja metszeteken. Ennek lényege a következő. A teljes sejt DNP-jének részét képező DNS-t denaturálják, majd radioaktív izotópokkal jelölt RNS-frakciókkal dolgozzák fel. Ennek eredményeként a DNS-en autoradiográfiás úton kimutathatóak azok a területek, amelyek komplementerek ezekkel az RNS-frakciókkal, vagyis ez utóbbiak transzkripciójának helyei, vagyis lehetővé válik bizonyos gének lokalizációjának pontos meghatározása.
A kísérleti módszeren belül a sejt egészének vagy egyes összetevőinek funkcionális szerveződését vizsgálják úgy, hogy külső hatások segítségével megváltoztatják állapotát. A sejt vagy alkotóelemei életaktivitásában bekövetkezett változások megfigyelésével következtetéseket vonhatunk le a vizsgált mechanizmusok bizonyos tulajdonságaira. Ez a fajta módszer mára igen elterjedt a citológia egyes részein, és egyes területein a sejtszerkezetek elemzésének citofiziológiai aspektusa még mindig domináns helyet foglal el.
Pontosan ez a probléma állapota a sejtfelszíni apparátus szállítási funkciójával kapcsolatban. Egyrészt jelentős előrelépés történt a kérdéskör vizsgálatában: a citofiziológiai elemzések eredményei alapján lehetőség nyílt az anyagok transzmembrán transzportjának típusainak azonosítására és a transzportrendszerek különböző tulajdonságainak jellemzésére. Másrészt a transzmembrán transzport mechanizmusai kérdésének végső megoldása csak akkor lehetséges, ha tisztázzuk a membránok lipid-protein rendszerének sajátos szerveződését, és pontosan ismerjük a membrán fennmaradó komponenseinek tulajdonságait és szerepét. membrán transzportrendszerek, azaz a plazmamembrán és a sejt teljes felületi apparátusának szerkezeti-biokémiai szintű elemzése.
A transzmembrán transzport citofiziológiai vizsgálatának korlátozott lehetőségeit egyértelműen bizonyítja az ioncsatornák szerveződésének kérdésének állása, amelyek számos fontos folyamatban, így például az idegimpulzus terjedésében is nagy szerepet játszanak. Különböző citofiziológiai módszerek egész arzenáljával kimutatták, hogy a plazmamembránban speciális csatornák vannak a Na, K, Cl ionok számára, amelyek tulajdonságaik különböznek egymástól. Szerkezeti felépítésükre vonatkozó konkrét ismeretek azonban egyelőre a fehérjetermészetükre vonatkozó közvetett adatokra korlátozódnak. Így különösen az ioncsatornák és általában a membrántranszportrendszerek szerveződésének kérdésének megoldása nyilvánvalóan a szerkezeti és biokémiai módszerekben jártas tudósok kezébe kerül, mivel ebben az esetben számos és nagyon értékes tényt nyernek a citofiziológiai tanulmányok csak az első fenomenológiai szakaszt képviselik ezen általános sejtmechanizmusok elemzésében. Ennek ellenére a sejtek tanulmányozásának bizonyos vonatkozásaiban a citofiziológiai megközelítés sokat segíthet.
Jelenleg a citofiziológiai kutatási technikák sokféleségét meghatározza egyrészt a citológusok rendelkezésére álló szerek egyre bővülő arzenálja, másrészt az ezeknek a szereknek a sejtre gyakorolt hatása következtében fellépő változások elemzésére szolgáló finom módszerek alkalmazása. Korábban a sejtek külső ágensek hatására bekövetkező változásainak elemzésére a fiziológusok által ismert módszereket alkalmaztak, mint például az elektromos potenciálok rögzítése, a sejtlégzés oxigénabszorpcióval történő felmérése, a festékszorpció kvantitatív értékelése, a sejtfestődés minőségi változásainak rögzítése stb. majd ma már, hasonló célokra egyre inkább a szerkezeti-funkcionális irányra jellemző módszereket alkalmazzák: ultrastrukturális változások elektronmikroszkópos vizsgálata, szintetikus folyamatok autoradiográfiás elemzése stb.
A kísérleti vizsgálatok során használt szerek között két fő csoport különíthető el. Az első csoportba azok az anyagok tartoznak, amelyeknek a sejten belüli „alkalmazási pontja” többé-kevésbé ismert – ezek olyan anyagok, amelyek blokkolják az intracelluláris metabolizmus egyes részeit (például aktinomicin D, amely gátolja a transzkripciót, vagy puromicin, amely blokkolja a fehérjeszintézist, 2,4-dinitrofenol, amely szétválasztja a légzést és az oxidatív foszforilációt, bizonyos sejtszerkezeteket szelektíven elpusztító anyagok (például a mikrotubulusokat elpusztító kolhicin vagy a mikrofibrillákra ható citokalazin B). A második csoportot az úgynevezett komplex hatású szerek alkotják, amelyek általában megváltoztatják a sejtanyagcserét - hőmérsékletet, ozmózisnyomást, pH-t stb. számos kérdés a légzés és a foszforiláció összekapcsolásával kapcsolatban a mitokondriumok légzőláncában; az RNS és a fehérjeszintézis gátlók alkalmazása lehetővé tette a riboszómák és a transzkripciós folyamatok fehérjeszintézisének egyes részeinek tanulmányozását; kolchicin és citokalazin segítségével tisztáztuk a mikrotubulusok és mikrofilamentumok szerepét az intracelluláris transzport folyamataiban.
A második csoportba tartozó (komplex hatás) szerek előnye, hogy természetesebbek a sejtek számára, mivel a sejtek természetes körülmények között hasonló változásokat tapasztalnak a külső környezetben. Ugyanakkor a sejtanyagcsere szinte minden aspektusára hatással vannak, megnehezítve a bekövetkező változások elemzését. Mindazonáltal az ilyen szerek sejtre gyakorolt hatásának vizsgálata önálló jelentőségű, és feltétlenül szükséges a sejtek változó környezeti tényezőkhöz való alkalmazkodási mechanizmusainak tanulmányozásához, a sejtek reakciójában a specifikus és a nem specifikus folyamatok közötti kapcsolat kérdésének megoldásához. külső hatások és más hasonló feladatok, amelyek fontos szerepet játszanak a sejtintegráció problémájának kialakulásában.
A sejtek funkcionális szerveződésének vizsgálatában nagy jelentősége van az egyes sejtrendszerek közötti kölcsönhatási mechanizmusok elemzésének. Ez a probléma sok esetben speciális kísérleti modellek létrehozásával megoldható. Ennek legjellemzőbb példái a különböző objektumokban (protozoonok, kétéltű tojások) végzett nukleáris transzplantáció; szomatikus sejt hibridizáció; sejtrészek átültetése protozoonokból; számos egyéb mikrosebészeti technikát alkalmazó tanulmányok, amelyeket protozoológiai tárgyakon és in vitro tenyésztett emlőssejteken végeztek.
Ilyen modellek segítségével a legfontosabb általános citológiai kérdéseket tanulmányoztuk. Például a differenciált kétéltűsejtek magjának saját magtól mentes tojásba való átültetésével kapcsolatos kísérletek eredményei szolgáltatták az egyik legmeggyőzőbb érvet a differenciális génaktivitás elmélete mellett. Ez utóbbi lényege egy többsejtű szervezet differenciált sejtjei genomjainak szerkezeti azonosságának megállapítása. Ebből következik az az alapvetően fontos feltevés, hogy a differenciálódás folyamata nem a sejtek örökletes apparátusának visszafordíthatatlan változásai révén megy végbe, hanem egy olyan génkészlet aktivitásának szabályozása révén, amely egy adott szervezet minden sejtjére azonos.
Nagyon érdekes tényeket fedeztek fel egy kísérleti modellben egy hibrid sejt - egy csirke eritrocita és egy emlős rákos sejt - dedifferenciálódási folyamatának tanulmányozására. Ennek a heterokarionnak az egyedisége abban rejlik, hogy amikor egy csirke vörösvértest egyesül egy rákos sejttel, hemoglobin hemolízis megy végbe, és a normál, szinte teljesen inaktivált vörösvértest mag a rákos sejt citoplazmájába kerül. Így itt egy differenciált sejtmagot ültetnek át az aktív citoplazma szokatlan körülményei közé. Ezen magok szerkezeti felépítésében bekövetkezett változások gondos megfigyelése azt mutatta, hogy új körülmények között jelentősen megnövekszik a térfogatuk. A citoplazmából származó fehérjék jelentős szerepet játszanak a nukleáris duzzanatban. Ezek a külső változások az eritrocita nukleáris apparátusában a belső szervezetének mélyreható átstrukturálási folyamatait tükrözik, amelyek a „csirke” hírvivő RNS-ek transzkripciójának újraindulását eredményezik. A benne található információk „csirke” fehérjék szintézise formájában való megvalósítása azonban nem történik meg addig, amíg a csirke eritrociták nukleáris apparátusában nukleolus nem képződik, és meg nem kezdődik a riboszómális RNS szintézise. Így a kísérleti modellek alapos elemzése azt mutatta, hogy a nukleáris apparátus aktivitása felett összetett citoplazmatikus szabályozás áll fenn.
Kísérleti modellek segítségével számos egyéb fontos általános citológiai kérdés megoldására volt lehetőség. Például az anafázis kromoszómák mozgásmechanizmusainak kérdését sikeresen tanulmányozták a natív mitotikus apparátuson, amelyet egy tengeri sün fragmentáló blasztomerjeiből izoláltak és a sejten kívül működtek. Főleg kísérleti modellek felhasználásával sikerült megállapítani a sejtszerveződés egy széles körben elterjedt általános mintáját, nevezetesen a szigorú ok-okozati elv hiányát az összetett intracelluláris folyamatok összekapcsolásában. Kiderült, hogy az olyan többkomponensű folyamatok, mint a sejtreprodukció, a szintézis és a nagy polimertartalmú vegyületek intracelluláris transzportja stb., különálló, viszonylag autonóm szakaszokból állnak, amelyeket nem köt össze szoros ok-okozati kapcsolat. Ennek a mintának a tisztázása egyrészt megteremti az előfeltételeket a sejtszerveződés elképesztő plaszticitásának mechanizmusainak megértéséhez. Másrészt ugyanez a minta az alapja az ilyen folyamatok integrálódási mechanizmusainak tanulmányozásának egy teljes sejtrendszerben normál körülmények között.
Jelenleg növekszik azoknak a kísérleti modelleknek a száma és változatossága, amelyeket bizonyos specifikus általános citológiai problémák megoldására terveztek. Ez jelentősen hozzájárul ismereteink fejlődéséhez a citológia viszonylag kevéssé tanulmányozott területén - a szubcelluláris rendszerek interakciós és integrációs mechanizmusaiban.
Hangsúlyozni kell, hogy a sejtszerveződési mintázatok vizsgálatának kísérleti megközelítése keretében végzett kutatások sajátossága azon kritériumok és jellemzők egyre mélyebbé tétele, amelyek alapján az egyes sejtstruktúrák integrációs mechanizmusait és sajátos funkcióit elemzik. Egyúttal világossá válik, hogy az ilyen jellegű kutatások előtt álló problémák sikeres megoldása csak a strukturális-funkcionális megközelítés módszereinek széles körű gyakorlati bevezetésével lehetséges.
Az általános citológiai kutatás összehasonlító citológiai módszerének lényege, hogy tisztázza a sejtszerveződés általános mintázatait, felhasználva az élő természet által a tudós számára biztosított változatosságot. Az összehasonlító módszernek két aspektusa van. Egyrészt hagyományosan az egyes sejttípusok közötti rokon kapcsolatok azonosítására használják (különösen az egysejtű szervezetek esetében). A prokarióta, valamint az alacsonyabb és magasabb rendű eukarióta sejtek így létrejött és finom citológiai kritériumok alapján végzett filogenetikai szisztematikája alapján nyomon követhetővé válik mind az egyéni privát sejtrendszerek kialakulásának, mind a sejt, mint a sejt, mint a sejt, a szabályozásának és integrációjának általános mechanizmusai. Integrál rendszer Példaként Az összehasonlító citológiai elemzésnek ez a fajta alkalmazása a sejtkutatásban érdekes adatokkal szolgálhat a prokarióta, alacsonyabb és magasabb rendű eukarióta sejtek sejtmagrendszerének finom szerveződéséről.
Az eukarióta sejtek nukleáris apparátusának felépítésének alapvető jellemzői a komplex nukleáris felszíni apparátus jelenléte, a kromoszómákban a prokarióta sejtekhez képest jóval nagyobb mennyiségű kromoszómák koncentrálódása, és végül az egyedi DNS-csomagolás az alapelemek segítségével. fehérjék - hisztonok Az alsóbbrendű eukarióták nukleáris apparátusának összehasonlító citológiai elemzése lehetővé tette közöttük olyan sejtek azonosítását, amelyek magszerkezetüket tekintve köztes helyet foglalnak el a pro- és eukarióta sejtek között. A páncélos flagellátok tipikus nukleáris felszíni apparátussal rendelkeznek, de kromoszómáikat, akárcsak a prokarióták esetében, körkörös DNS-molekulák alkotják, amelyek hisztonok részvétele nélkül, minden eukariótára jellemző kompakt struktúrákba szerveződnek.
A közelmúltban a pro- és eukarióta sejtek genomjának szerveződésének alapvető jellemzőinek felfedezése kapcsán fontossá vált ezekben a szervezetekben, valamint a mezokariótákban és az alacsonyabb rendű eukarióták sejtjeiben zajló transzkripciós és RNS-érési folyamatok összehasonlítása. Az ilyen összehasonlítások eredményeként elképzelhető, hogy jelentős változások történtek a fő organizmuscsoportok családi kapcsolatairól alkotott hagyományos elképzeléseinken, és különösen a pro- és eukarióta sejtek közötti kapcsolatokban.
A citológiai problémák evolúciós megközelítésének hagyományos alkalmazásának második példája lehet egy hipotézis felállítása a kromoszómák evolúciós folyamatában a kromoszómák egyenlő eloszlásának mechanizmusaira vonatkozóan, amelyet egy összehasonlító elemzés alapján dolgoztak ki. a kromoszóma-divergencia számos változata protozoákban és alacsonyabb rendű növényekben. Ezekben az esetekben a magmembrán membránok aktívan részt vesznek a kromoszóma szegregáció folyamataiban, ami lehetővé teszi bizonyos homológiát a prokarióta sejtekkel, amelyben a sejtmembrán vezető szerepet játszik a testvérkromoszómák leánysejtek közötti egyenletes eloszlásában.
Végül az általános citológiai problémák hagyományos evolúciós megközelítésének harmadik példája lehet a mitokondriumok és a kloroplasztiszok eredetének széles körben elterjedt szimbiotikus hipotézise. Lényege abban a feltevésben rejlik, hogy ezek az energia-anyagcsere fontos szervei olyan prokarióta organizmusokból származnak, amelyek az eukarióták evolúciójának viszonylag korai szakaszában támadták meg az eukarióta sejteket.
Annak ellenére, hogy az ilyen általános biológiai konstrukciók fontosak az általános citológia fejlődésében, az általános citológiai problémák kialakulásának hagyományos történeti megközelítése ma még meglehetősen korlátozottan használható. Ennek a helyzetnek az egyik fő oka az evolúciós folyamat sajátos és még nem kellően tanulmányozott sajátosságainak jelenléte a sejtes és szubcelluláris szerveződési szinteken, ami rendkívül megnehezíti az egysejtű szervezetek egyes csoportjai közötti rokon kapcsolatok meghatározását, és ennek következtében. , megalapozott evolúciós új hipotézisek felállítására az általános citológia területén.
Jelenleg az összehasonlító citológiai módszer alkalmazásának egy másik aspektusa is elterjedtebb, amely nem azt a célt követi, hogy egy adott sejtszerkezet vagy folyamat történeti feltételrendszerét közvetlenül megvilágítsa. A modern általános citológiában az összehasonlító módszer alkalmazásának ezen aspektusa számos módosításon ment keresztül.
Az első szakaszban, az alapvetően új morfobiokémiai módszerek citológiai analízis gyakorlatba való bevezetésének időszakában, a vizsgálat tárgyának megválasztását a következő szempontok határozták meg. Először is, egy adott objektum kényelme volt fontos az alkalmazott módszer alkalmazásához. Másodszor, ennek a jelnek a kifejeződési foka a vizsgált sejtben fontos szerepet játszott. Így az eukarióta sejtek szerveződésének általános törvényszerűségeinek tanulmányozására az emlős májsejtek harmonikusan fejlett membránszervecskéivel a hasnyálmirigy-sejteken végzett klasszikus munka az intracelluláris transzport és a váladékérés folyamatainak elemzésével. és nyálkahártya serlegsejtes emlősök.
Az Escherichia colit széles körben alkalmazzák a prokarióta sejtek szerveződésének komplex citológiai és molekuláris biológiai vizsgálataiban; Az alacsonyabb rendű eukarióták szerveződésének tanulmányozására szolgáló modellek az élesztő és a penész voltak. Kiderült, hogy az ezeken az objektumokon kialakult minták univerzális jelentőséggel bírnak, mivel sok esetben alapvetően hasonlóak minden eukarióta vagy minden prokarióta sejtben. Sőt, a szubcelluláris szerveződés számos mintázata, különösen molekuláris és szupramolekuláris szinten, univerzálisnak bizonyult mind a pro-, mind az eukarióta típusú sejtek esetében (membránok szerveződése, riboszóma szerkezetének elve stb.), annak ellenére, hogy hogy a megnevezett sejttípusok bizonyos tekintetben alapvetően különböznek egymástól. Ez a körülmény adta az ötletet, hogy az objektumok korlátozott körén, módszertani szempontból kényelmesen lehet alapvető általános citológiai problémákat kidolgozni, majd a kialakult mintázatokat kiterjeszteni más sejtekre, alapvetően hasonló szerveződésük miatt.
Az utóbbi években azonban az összehasonlító megközelítésnek ezt az egyszerűsített alkalmazását bírálni kezdték, mivel a modern citológiai módszereket bevezetik a speciális sejtbiológiai tudományokba - speciális citológia, protozoológia, alsóbbrendű növények botanikája. Az ezeken a tudományterületeken alkalmazott morfobiokémiai elemzés lehetővé tette olyan tények megállapítását, amelyek a sejtszerveződés egyik vagy másik általános jellemzőjének specifikus megvalósításában óriási sokféleséget jeleznek, sokkal nagyobb diverzitást, mint ami a korábban „modell” objektumokon kapott eredményekből következett. Ez a sokféleség különösen nagy a sejtszerveződés legmagasabb alrendszeri és szisztémás szintjein. Jellemző az olyan összetett és többkomponensű folyamatokra is, mint az intracelluláris anyagcsere és transzport folyamatai, vagy a genetikai anyag egyformán örökletes eloszlása a sejtosztódás során.
A modern módszertani képességek szintjén nyert nagy mennyiségű összehasonlító citológiai anyag általánosítása arra késztetett bennünket, hogy feladjuk az összehasonlító módszer szerepének fent említett leegyszerűsített elképzelését. Ebben a tekintetben az általános citológiában (különösen az eukarióta sejtekkel kapcsolatban) a domináns pozíciót az a gondolat szerezi meg, hogy összehasonlító módszert kell alkalmazni az egyes sejtrendszerek vagy a funkcionális aktivitásukban hasonló folyamatok elemzésére. megnyilvánulásaik sokfélesége meghatározott sejtekben Ezzel a megközelítéssel a különleges érthetőséget nem a „tipikus”, „átlagos” sejtek okozzák, hanem éppen ellenkezőleg, az átlagos szerveződéstípustól élesen eltérő sejtek. amelyek bizonyos jellemzői hipertrófiásak.
A legtöbb ilyen „kikerülő” variáns a magasabb rendű többsejtű szervezetek sejtjei között található, ahol az egyes szövetrendszereken belül a sejtek messzemenő specializációja fejlődik ki. Az átlagos típustól való „kijátszás” esetei széles körben elterjedtek a magasabb protozoonok körében is, amelyek evolúción mentek keresztül, miközben megtartották az egysejtű szerveződési szintet. Az ilyen típusú atipikus sejtek vizsgálata során számos új érdekes tényt sikerült azonosítani, amelyek jelentősen elmélyítik mind a sejtszerveződés általános törvényszerűségeinek, mind pedig a sejtrendszerek megfigyelt sokféleségét meghatározó evolúciós plaszticitásnak a megértését. . Ugyanakkor, amint azt fentebb megjegyeztük, a speciális tudományok esetében a legnagyobb érdeklődést az összes sejtre jellemző közös jellemzők különböző objektumokban való megnyilvánulásának sajátossága okozza.
A speciális tudományokkal ellentétben általános citológiai megközelítéssel ez a kérdés egy kicsit más síkon kerül fel, mert a kutató arra törekszik, hogy egy adott tulajdonság konkrét megnyilvánulásai mennyire elterjedtek a különböző sejtekben, milyen kombinációja az általános mechanizmusoknak, ill. pontosan mi okozza. Például a protozoológusoknak sikerült felfedezniük a macronucleus kialakulásának nagyon érdekes dinamikáját a konjugáció után gyomorciliáris csillótestekben. A kialakuló makronukleuszban jelentősen megnő a DNS mennyisége, majd az örökítőanyag éles csökkenése figyelhető meg (akár 93%). Ez a genetikai anyag redukciós folyamata számos többsejtű állatcsoport (egyes rovarok, fonálférgek) szomatikus sejtjeiben is végbemegy. A fennmaradó, kis mennyiségű, de a makronukleusz működéséhez szükséges összes információt tartalmazó DNS sokszor replikálódik. Ennek eredményeként egy definitív makronukleusz jön létre, amely nemcsak a DNS mennyiségében, hanem minőségi összetételében is különbözik a mikronukleusztól. A nem működő gének túlnyomó többsége itt hiányzik, míg a működő lókuszokat jelentős számú kópia képviseli.
Ezek a tények éppen azért nagy általános citológiai érdeklődésre tartanak számot, mert az itt megfigyelt jelenségek általában nem egyszerűen csak a magasabb egysejtű szervezetekre jellemző paradox jelekként működnek. Így a politenizációs folyamatok, a kromoszómális DNS egyes szakaszainak szelektív replikációja, és végül a genom jelentős szakaszainak szelektív redukálása - mindezek a jelenségek a többsejtű szervezetek speciális sejtjeiben is előfordulnak. Valószínűleg közös elemi mechanizmusok alapján hajtják végre. A gasztrociliáris csillókban a makronukleusz kialakulása során a nukleáris berendezésben végbemenő változások összetett folyamatának sajátossága pedig elsősorban az eukarióta sejtek közös, univerzális elemi mechanizmusainak sajátos kombinációjának köszönhető. Ez a fajta elképzelés manapság kezd széles körben elterjedni az általános citológiában. Rendkívül ösztönzik a célzott, összehasonlító citológiai vizsgálatokat, amelyek célja a fontos általános citológiai problémák feltárása. Anyag az oldalról
Célzott, összehasonlító citológiai vizsgálatra példa az eukariótákban a mitózis során a kromoszómák egyenlő eloszlásának mechanizmusainak vizsgálata különböző kovaalmafajok mitózisának elemzésével: ezeken a tárgyakon a metazoa sejtek tipikus mitózisaival ellentétben lehetőség van egyértelműen morfológiailag nyomon követheti a mikrotubulus-szervező központok komplex változásait, a mikrotubulus félorsók kialakulását és kölcsönös divergenciáját, a kromoszómák divergenciáját a sejt pólusaihoz egy sajátos szerkezet metafázisban - gallér - kialakulása révén.
A tubulin-dynein mechanokémiai rendszernek a metazoa sejtekben a kromoszómák anafázisos mozgásában betöltött vezető szerepére vonatkozó legfrissebb adatok fényében nagyon valószínű, hogy ez a rendszer a kovamoszatokban is jelen van, vagyis itt is csak egy az összes sejtre jellemző elemi mechanizmusok sajátos kombinációja, amelyek meghatározzák a mitózis során lezajló mechanokémiai folyamatokat.
Nyilvánvalóan e mechanizmusok elemzéséhez, amelyek feltárása az általános citológia egyik nagyon sürgető problémáját jelenti, ígéretes lenne egy olyan tárgy, ahol egyértelműen megkülönböztethetők és morfológiailag kifejeződnek.
Az ilyen jellegű példák száma folyamatosan növekszik. Ez egyrészt annak köszönhető, hogy a komplex, modern módszerek egyre szélesebb körben bekerülnek az egyes citológiai, protozoológiai és botanikai kutatások gyakorlatába, másrészt pedig annak, hogy az összehasonlító citológiai vizsgálatok során egyre inkább felhalmozódnak a tények. fontosak az általános citológia szempontjából, és figyelmének középpontjában állnak. Mindez pedig oda vezet, hogy az összehasonlító citológiai elemzés rendkívül fontos helyet foglal el a citológiában.
A modern általános citológiai kutatás fő irányainak és szempontjainak rövid ismertetése azt mutatja, hogy a citológia fejlődésének ebben a szakaszában az egyes irányok meglehetősen világos körülhatárolása és szintézise egyaránt megvan. A megkülönböztetés mind módszertani szempontból, mind pedig az egyes irányok és megközelítések által felvetett konkrét problémák megoldási logikája tekintetében történik. A citológiai vizsgálatok morfofunkcionális aspektusát a sejtszerkezetek elemzésének diszkrét megközelítése uralja. A sejtszerveződési mintázatok vizsgálatának kísérleti megközelítésének egyik legfontosabb jellemzője, hogy a sejtrendszerek és a teljes sejt szerveződésének általános integrációs mechanizmusainak elemzésére összpontosít. Ugyanakkor, mint fentebb már hangsúlyoztuk, az ilyen vizsgálatok előtt álló problémák megoldása lehetetlen a morfofunkcionális megközelítésben rejlő módszerek széles körű alkalmazása nélkül. A kísérleti elemzés fenomenológiai jellemzőt ad bizonyos sejtmechanizmusok és intracelluláris folyamatok tulajdonságaira, megteremtve ezzel a szükséges alapot a szerkezeti és biokémiai módszerek gazdag arzenáljának alkalmazásához.
Így az általános citológia fejlődésének jelenlegi szakaszában megvannak az előfeltételei a citológiai kutatás e két aspektusának nagyon szoros kombinációjának. Ez természetes, hiszen végső soron mindkét megközelítés ugyanazt a célt követi - a sejtszerkezetek funkcionális szerveződésének és a folyamatok szabályozási mechanizmusainak feltárását az egész sejtrendszerben.
Az általános citológiai problémák elemzésének összehasonlító citológiai megközelítése különleges helyet foglal el a modern általános citológiában. Az összehasonlító citológiai analízis morfofunkcionális és kísérleti megközelítések alapján nyert adatok alapján történik, azaz módszertanilag a citológiai kutatás minden fő szempontja szorosan összefügg egymással.
Az összehasonlító citológiai megközelítés sajátossága, a speciális helyzetét meghatározó specifitás az élő természet különböző objektumainak célirányos felhasználása az egyes sejtstruktúrák általános szerveződési mintáinak, az intracelluláris folyamatoknak és az integrációs mechanizmusoknak a megnyilvánulási formáinak sokféleségében történő tanulmányozására. különböző típusú sejtekben.
Tehát, amint az a fentiekből látható, a citológiai kutatások fő irányai nagymértékben meghatározzák az általános citológia modern fejlődési szakaszának sajátosságait, és meghatározzák annak szoros kapcsolatát a kapcsolódó biológiai tudományokkal. Ennek a szakasznak az egyik legjellemzőbb sajátossága a citológiai kutatások módszertani értelemben vett legfontosabb területeinek szoros kapcsolata. Ezenkívül az ilyen módszertani integráció gyakran túlmutat az általános citológia keretein.
A citológiai munkában széles körben alkalmazzák a tisztán biokémiai és molekuláris biológiai módszereket, és fordítva, a biokémiai és molekuláris biológiai vizsgálatokban a citológiai morfológiai módszereket. A kapcsolódó tudományok módszertani integrációja és végső céljaik egysége egy új szintetikus sejttudomány - sejtbiológia - kialakulásához vezetett. Egyesíti a citológiát, a szerkezeti biokémiát, a molekuláris biológiát, a molekuláris genetikát és a speciális biológiai tudományokat a sejtszintű szerveződésről. A rokon tudományok ilyen kombinációja kétségtelenül progresszív jelenség. Az ilyen szintézis ellenére azonban a tudományok mindegyike megőrzi saját módszertani sajátosságait és specifikusságát a sejtszerveződési problémák megfogalmazásában és fejlesztési módszereiben. Ebben a szintetikus tudományban jelenleg a molekuláris biológiai és molekuláris genetikai kutatások dominálnak. Ez a helyzet a sejtszerveződési alacsonyabb szintekkel kapcsolatos ismereteink rohamos fejlődésének köszönhető, de ez csak átmeneti jelenség.
Valójában a sejt új szintetikus tudományában a vezető helyet az általános citológiának kell elfoglalnia - az élő anyag sejtszintű szerveződésének általános törvényszerűségeinek tudományának. Az élőanyag ilyen szerveződési szintjével foglalkozó modern biológia tudományok közül az általános citológia, a célzott, szerkezeti és biokémiai módszerekre épülő összehasonlító citológiai megközelítést bővítve, a legfelkészültebb a hatalmas tényanyag mélyreható általános biológiai általánosítására, az élőanyag diszkrét elemzésével. - sejtszerkezetek számos sejttípusban. Az általános citológiának vezető szerepet kell betöltenie az általános sejtintegrációs mechanizmusok elemzésében. Ennek fontos előfeltétele az új kísérleti modellek gyors fejlődése. Mélyreható elemzésük modern módszerekkel és a kísérleti modellek széles körű bevezetése a célzott összehasonlító citológiai vizsgálatokba előrelépést biztosít a sejtszerveződés egyik fő problémájának - a sejtintegráció problémájának - megoldásában.
Mivel tényanyag gyűlik fel a sejtintegráció elemi univerzális mechanizmusairól és azok módosulásának hatóköréről, az általános citológia az a feladat, hogy mélyrehatóan elemezze az egyes sejtrendszerek szerveződésének történeti feltételrendszerét és általában a sejtszerveződést. valamint az evolúciós folyamat sajátosságai az élő anyag sejt- és szubcelluláris szerveződési szintjén. A probléma megoldását megkönnyíti az általános citológiai vizsgálatokban ma már jól látható tendencia, hogy egyesítik a sejt egyes komponenseinek diszkrét elemzését a sejt kg-ának vizsgálatával. egy teljes rendszerhez.
Ezen az oldalon a következő témákban található anyagok:
Az életszervezés sejtszintű
16. § A sejtek tanulmányozásának története. A citológiai kutatás módszerei.
A sejtek tanulmányozásának története.
A sejtek világa egészen a 17. század közepéig teljesen ismeretlen maradt, amikor az emberek megtanulták a lencséket csiszolni és a látás javítására használni.
A mikroszkóp egyik első alkotója volt Robert Hooke fizikus, meteorológus, biológus, mérnök, építész. IN 1665"makrográfia" néven rajzalbumot adott ki, amely mikroszkóp alatt mutatta be megfigyeléseit.
Hooke egyik tehetséges kortársa a holland volt Anthony van Leeuwenhoek, aki 200 saját tervezésű mikroszkópot készített. Leeuwenhoek 270-szeresére növelte a tárgyak számát, és kiemelkedő felfedezéseket tett.
Robert Brown 1833-ban felfedezte a sejtmagot egy sejtben. Után 1825 Jan Purkinje hatékony módszereket dolgozott ki a mikroszkopikus berendezések preparátumainak elkészítésére és festésére.
A növények számára javasolt sejtelmélet 1837 német botanikus Matthias Schleiden,és barátja, a fiziológus kiterjesztette az állatvilágra Theodor Schwann. Kicsit később kiegészítették Rudolf Virchow, amely be 1885 megfogalmazta a tételt: „Minden sejt sejtből származik”.
század közepén. A sejtelmélet általánosan elfogadottá vált és a sejttudomány alapjává vált. citológia. A 19. század végére. sejt számos összetevőjét fedezték fel. A tudósok leírták és elnevezték őket.
De be 1945 A citológusok először vizsgáltak sejteket elektronmikroszkóp segítségével, és számos korábban ismeretlen struktúrát láttak. Tehát a citológia fejlődésében a döntő szerep más tudományok, különösen a fizika új felfedezéseié.
A citológiai kutatás módszerei.
A fő módszer az fénymikroszkópos módszer. Fénymikroszkóp használatával jár, de csak speciálisan elkészített citológiai készítmények vizsgálhatók fénymikroszkóp alatt.
A készítmények elkészítéséhez a citológusok tárgylemezeket és speciálisan előkészített vizsgálható tárgyakat használnak.
Leggyakrabban ezek a szerkezetek színtelenek, ezért speciális festékekkel kell festeni, minden alkalommal más és más, attól függően, hogy milyen szerkezeteket szeretne látni.
Két módszer létezik: nyomás alatti preparátumok készítésének módszere - a vizsgált tárgyat egyszerűen egy rétegre zúzzák egy tárgylemez és egy fedőüveg közé, valamint egy eljárás vékony, egyetlen sejtrétegből álló metszetek elkészítésére.
Élő sejtek tanulmányozására használják fáziskontraszt mikroszkópos módszer. Azon alapul, hogy egy átlátszó cella egyes szakaszai sűrűségben és fénytörésben különböznek egymástól.
Élő sejtek tanulmányozásakor azt is használják fluoreszcens mikroszkópos módszer. Jelentése abban rejlik, hogy számos anyag fényenergiát nyelve képes izzani. Például, ha a növényi sejteket fluoreszcens mikroszkópon keresztül nézi, akkor a sötétkék testen vörös szemcséket fog látni, amelyek élénken világítanak - ezek kloroplasztiszok.
Van egy módszer, amely jelölt izotópokat használ - autoradiográfiás módszer- izotópokkal jelölt anyagok regisztrációja. Ezzel a módszerrel láthatja, hogy a sejt mely részei kapnak radioaktív izotópokkal jelölt anyagokat.
Elektronmikroszkópos módszer A citológus felfedezte azokat a sejtstruktúrákat, amelyek mérete kisebb, mint a fény hullámhossza. Ennek a módszernek köszönhetően lehetővé vált olyan vírusok és organellumok vizsgálata, amelyeken fehérjeszintézis megy végbe (riboszómák).
A citológusok a sejtek különféle összetevőit is beszerezhetik és tanulmányozhatják sejtfrakcionálás. A sejtet először megsemmisítik, majd egy speciális eszközzel - centrifugával - izolálják a sejtszerkezeteket.
Sejtkultúra felhasználási módja az emberi, állati vagy növényi testből izolált sejtek, szövetek, kis szervek vagy azok részei hosszú távú tárolásának és tenyésztésének módszere speciális tápközegben. Ennek a módszernek egy fontos előnye, hogy mikroszkóppal megfigyelhető a sejtek létfontosságú tevékenysége.
A citológiai módszerek jelentősége az emberi betegségek diagnosztizálásában és kezelésében.
1) Citológiai módszerek az orvostudományban az emberi test élettani állapotának tanulmányozására használják a sejtek szerkezetének tanulmányozása alapján. Vérbetegségek azonosítására, rosszindulatú és jóindulatú daganatok felismerésére, számos légúti, emésztési, vizeletürítési, idegrendszeri megbetegedésre és ezek kezelésére szolgálnak.
2) Őssejt egy éretlen sejt, amely képes önmegújulásra és a test speciális sejtjévé fejlődni. A felnőtt szervezetben az őssejtek főként a csontvelőben találhatók, és nagyon kis mennyiségben minden szervben és szövetben. Számos betegség kezelésére használhatók.
17. § A prokarióta és eukarióta sejtek felépítése.
A sejtszerkezet egysége.
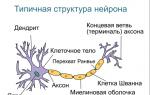
Bármely cella tartalmát egy speciális szerkezet választja el a külső környezettől - plazmamembrán (plazmalemma). Ez az elszigeteltség lehetővé teszi, hogy egy nagyon különleges környezetet hozzon létre a sejten belül, ellentétben azzal, ami körülveszi. Ezért olyan folyamatok fordulhatnak elő a sejtben, amelyek sehol máshol nem fordulnak elő életfolyamatokat.
Az élő sejt plazmamembránnal határolt belső környezetét ún citoplazma. Ez magában foglalja hialoplazma(átlátszó alapanyag) és sejtszervecskék, valamint különféle nem állandó szerkezetek - zárványok. Bármely sejtben jelen lévő organellumok is magukban foglalják riboszómák, hol történik fehérjeszintézis.
Az eukarióta sejtek szerkezete.
Eukarióták- Ezek olyan élőlények, amelyek sejtjei rendelkeznek maggal. Mag- ez az eukarióta sejtnek az az organellumája, amelyben a kromoszómákban rögzített örökletes információ tárolódik, és amelyből az örökletes információ átíródik. Kromoszóma egy fehérjékkel integrált DNS-molekula. A mag tartalmaz nucleolus- az a hely, ahol a fehérjeszintézisben részt vevő egyéb fontos organellumok keletkeznek, riboszómák. De a riboszómák csak a sejtmagban képződnek, és a citoplazmában dolgoznak (azaz fehérjét szintetizálnak). Egy részük szabadon található a citoplazmában, egy részük membránokhoz kötődik, hálózatot alkotva, amit ún. endoplazmatikus.
Riboszómák- nem membrán organellumok.
Endoplazmatikus retikulum membránnal határolt tubulusok hálózata. Két típusa van: sima és szemcsés. A riboszómák a szemcsés endoplazmatikus retikulum membránjain helyezkednek el, így a fehérjék szintetizálódnak és oda szállítódnak. A sima endoplazmatikus retikulum pedig a szénhidrátok és lipidek szintézisének és szállításának a helye. Nincsenek rajta riboszómák.
A fehérjék, szénhidrátok és zsírok szintézise energiát igényel, amelyet az eukarióta sejtben a sejt „energia állomásai” állítanak elő - mitokondriumok.
Mitokondriumok- kettős membrán organellumok, amelyekben a sejtlégzés folyamata megy végbe. A szerves vegyületek a mitokondriális membránokon oxidálódnak, és a kémiai energia speciális energiamolekulák formájában halmozódik fel (ATP).
A sejtben is van egy hely, ahol a szerves vegyületek felhalmozódhatnak, és ahonnan el tudnak szállítani - ez van Golgi készülék, lapos membrán zacskók rendszere. Részt vesz a fehérjék, lipidek és szénhidrátok szállításában. A Golgi-készülék az intracelluláris emésztéshez organellumokat is termel - lizoszómák.
Lizoszómák- az állati sejtekre jellemző egymembrán organellumok olyan enzimeket tartalmaznak, amelyek képesek lebontani a fehérjéket, szénhidrátokat, nukleinsavakat, lipideket.
Egy sejt tartalmazhat membránszerkezettel nem rendelkező organellumokat, például riboszómákat és citoszkeletont.
Citoszkeleton- ez a sejt mozgásszervi rendszere, magában foglalja a mikroszálakat, csillókat, flagellákat, a sejtközpontot, amely mikrotubulusokat és centriolákat termel.
Csak a növényi sejtekre jellemző organellumok vannak - plasztidok. Vannak: kloroplasztok, kromoplasztok és leukoplasztok. A fotoszintézis folyamata a kloroplasztiszokban megy végbe.
Növényi sejtekben is vakuolák- a sejt salakanyagai, amelyek a víz és a benne oldott vegyületek tározói. Az eukarióta szervezetek közé tartoznak a növények, állatok és gombák.
A prokarióta sejtek szerkezete.
Prokarióták- egysejtű szervezetek, amelyek sejtjei nem rendelkeznek maggal.
A prokarióta sejtek kis méretűek, és a genetikai anyagot körkörös DNS-molekula (nukleoid) formájában tárolják. A prokarióta szervezetek közé tartoznak a baktériumok és a cianobaktériumok, amelyeket korábban kék-zöld algáknak neveztek.
Ha az aerob légzés folyamata prokariótákban fordul elő, akkor ehhez a plazmamembrán speciális kiemelkedéseit használják - mezoszómák. Ha a baktériumok fotoszintetikusak, akkor a fotoszintézis folyamata a fotoszintetikus membránokon megy végbe - tilakoidok.
A prokariótákban a fehérjeszintézis a riboszómák. A prokarióta sejteknek kevés organellumjuk van.
Az eukarióta sejtek organellumainak eredetének hipotézisei.
A prokarióta sejtek korábban jelentek meg a Földön, mint az eukarióta sejtek.
1) szimbiotikus hipotézis elmagyarázza az eukarióta sejt egyes organellumainak - mitokondriumok és fotoszintetikus plasztidok - megjelenésének mechanizmusát.
2) Intussuscepciós hipotézis- azt állítja, hogy az eukarióta sejt eredete abból származik, hogy az ősi forma aerob prokarióta volt. A benne lévő organellumok a héj egyes részeinek invaginációja és leválása következtében keletkeztek, majd funkcionális specializálódást követett más organellumok sejtmagjába, mitokondriumaiba, kloroplasztiszaiba.
18. § Sejthártyák. Anyagok szállítása a membránokon keresztül. A sejt felszíni apparátusa, funkciói.
Sejtmembránok.
Biológiai membránok- ezek vékony, szomszédos molekulaméretű struktúrák, amelyek a sejtek felszínén és a szubcelluláris részeken helyezkednek el, valamint tubulusok és vezikulák, amelyek behatolnak a protoplazmába. A biológiai membránok feladata az ionok, cukrok, aminosavak és egyéb anyagcseretermékek szállításának szabályozása.
Bármely membrán alapja a foszfolipidek kettős rétege.
A bilipid réteg azonban nem egy kész membrán, hanem csak az alapja. A fehérjéket ún membránfehérjék. A membránfehérjék határozzák meg a membránok számos tulajdonságát. A szénhidrátok szintén a membránok részét képezik, és komplexeket képeznek fehérjékkel vagy lipidekkel. A membrán egy bilipid rétegből áll, amelyben fehérjemolekulák lebegnek (vagy rögzítve vannak), egyfajta mozaikot képezve benne.
A membrán szerkezete megfelel a funkcióinak: transzport, gát és receptor.
1) Sorompó funkció. A membrán egy gát, amely megakadályozza a különféle vegyi anyagok és egyéb szerek bejutását a sejtekbe.
2) Receptor funkciók. A membrán felületén számos receptor található, amelyek specifikus reakciókat tesznek lehetővé különféle anyagokkal.
3) Szállítási funkció. Az ionok és anyagok szállítása a membránon keresztül történik.
A sejt lefedésével és a környezettől való elválasztásával a biológiai membránok biztosítják a sejtek és organellumok integritását. Fenntartja a kálium-, nátrium-, klór- és egyéb ionok egyenetlen eloszlását a protoplazma és a környezet között.
A sejtben különösen fontos membrán az plasmalemma- felületi membrán. Gát, transzport, receptor, jelző funkciókat lát el.
Anyagok szállítása a membránokon keresztül.
Két aktív folyamat van: exocitózis és endocitózis.
Az anyagokat a sejtből távolítják el exocitózis- intracelluláris vezikulák fúziója a plazmamembránnal. Az anyagok ezen keresztül juthatnak be a sejtbe endocitózis. Az endocitózis folyamata során a plazmamembrán homorúakat képez és megnövekszik, amelyek aztán lehámlanak és hólyagokká vagy vakuolákká alakulnak.
Az endocitózisnak két típusa van:
- Pinocitózis- folyékony és oldott anyagok felszívása kis buborékok segítségével;
- Fagocitózis- nagy részecskék, például mikroorganizmusok vagy sejttörmelékek felszívódása.
Fagocitózis esetén nagy buborékok keletkeznek, amelyeket ún vakuolák.
A molekulák membránokon haladnak át a folyamatokon: egyszerű diffúzió, könnyített diffúzió, aktív transzport.
Egyszerű diffúzió- Ez egy példa a passzív transzportra, amely nagyobb molekulakoncentrációjú területről alacsonyabb koncentrációjú területre halad át. Egyszerű diffúzióval lipidekben és kis töltés nélküli molekulákban (például vízben) oldódó nempoláris (hidrofób) anyagok behatolnak a sejtbe. A legtöbb anyag azonban a membránon keresztül jut el a membránba ágyazott transzportfehérjék segítségével. A címzésnek két formája van: megkönnyítette a diffúziót és az aktív transzportot.
Könnyített diffúzió koncentráció gradiens határozza meg, és a molekulák ennek a gradiensnek megfelelően mozognak. A molekula azonban feltöltött, szállítását a koncentráció gradiens és a membránpotenciál egyaránt befolyásolja.
Aktív közlekedés az oldott anyagok koncentrációgradiens ellenében történő transzportja az ATP energiájának felhasználásával. Az energiára azért van szükség, mert az anyagnak a diffúzió általi természetes mozgási hajlamával ellentétben az ellenkező irányba kell mozognia. Ilyen például a nátrium-kálium szivattyú. A diffúzió törvényei szerint a Na-ionok folyamatosan bejutnak a sejtbe, a K+-ionok pedig kikerülnek a sejtből. Ezen ionok szükséges koncentrációjának megsértése sejthalálhoz vezet.
A cella felületi berendezése.
A különböző prokarióta és eukarióta sejtek részekből állnak: felszíni berendezés, citoplazma, nukleáris készülék.
Felületi készülékek A sejtek három olyan funkciót látnak el, amelyek minden típusú sejt számára univerzálisak: gát, transzport, receptor. Számos specifikus funkciót is elláthat (például növényi sejtekben a sejtfal mechanikus turgor funkciója). A sejtek felszíni apparátusa rendszerekből áll: a plazmamembránból, a szupramembrán komplexumból és a szubmembránból (azaz szubmembránból) a mozgásszervi apparátusból.
plazmamembrán, vagy plasmalemma, a felszíni apparátus fő rendszere, minden sejt számára univerzális. Alatta egy szubmembrán rendszer található, amely a transzmembrán transzportban és befogadásban vesz részt, és a citoplazma része.
Szupramembrán szerkezet A felületi berendezés kölcsönhatásba lép a sejtek és a külső környezet vagy más sejtekkel. Állati sejtekben a szupramembrán komplex, ill glikokalix, fontos szerepet játszik a sejtek receptor funkciójában. A glikokalix szénhidrátokból áll, és viszonylag vékony és rugalmas.
A származtatott membrán feletti struktúrákhoz tartozik sejtfal. A növények, gombák és baktériumok sejtjeinek kell termelniük. A növények sejtfala cellulózt, gombákat - kitint, baktériumokat - mureint tartalmaz. Elég merev és nem zsugorodik. Víz, só és számos szerves anyag molekulája áthalad a sejtfalon. A plazmolízis és deplazmolízis jelensége növényi sejtekben.
Plazmolízis- ez a citoplazma elválasztása a membrántól, amikor a sejt hipertóniás, azaz hipertóniás állapotba kerül. kívülről koncentrált, oldat. Ha az állati sejteket hipertóniás oldatba merítjük, azok összezsugorodnak. Néha a plazmolizált sejtek életben maradnak. Ha az ilyen sejteket olyan vízbe merítjük, amelyben a sók koncentrációja alacsonyabb, mint a sejtben, deplazmolízis megy végbe.
Deplazmolízis- ez a növényi sejtek citoplazmájának visszatérése a plazmolízis állapotából az eredeti állapotba.
 Galileo-Galilei (1564-1642) (olasz filozófus, matematikus, fizikus és csillagász, aki jelentős hatással volt korának tudományára; a mikroszkóp feltalálója) Az egyik első mikroszkóp (1876)
Galileo-Galilei (1564-1642) (olasz filozófus, matematikus, fizikus és csillagász, aki jelentős hatással volt korának tudományára; a mikroszkóp feltalálója) Az egyik első mikroszkóp (1876)

 Fénymikroszkóp Robert Hooke (1635-1703) 1665 – „Mikrográfia” monográfia, amely leírja mikroszkópos és teleszkópos megfigyeléseit
Fénymikroszkóp Robert Hooke (1635-1703) 1665 – „Mikrográfia” monográfia, amely leírja mikroszkópos és teleszkópos megfigyeléseit



 A MIKROSZKÓP KÉSZÜLÉKE ÉS MŰKÖDÉSI ELVE Modern fénymikroszkóp 1. Mechanikai rész 1. 1. Test 1. 2. Mechanikai (minta) szakasz 1. 3. Távcsőrögzítés 1. 4. Fókuszáló mechanizmus 2. Fényrendszer 2. 1. Fényrendszer forrás 2. 2. gyűjtő 2. 3. kondenzátor 3. optikai rész 3. 1. objektívek 3. 2. szemlencsék
A MIKROSZKÓP KÉSZÜLÉKE ÉS MŰKÖDÉSI ELVE Modern fénymikroszkóp 1. Mechanikai rész 1. 1. Test 1. 2. Mechanikai (minta) szakasz 1. 3. Távcsőrögzítés 1. 4. Fókuszáló mechanizmus 2. Fényrendszer 2. 1. Fényrendszer forrás 2. 2. gyűjtő 2. 3. kondenzátor 3. optikai rész 3. 1. objektívek 3. 2. szemlencsék

 Sugarak útja szabványos mikroszkóp fényforrás kondenzátor minta lencse szemlencse szem Sugarak útja modern mikroszkópban fényforrás mintagyűjtő kondenzátor szemlencse p képe a mintáról. A MIKROSZKÓP KÉSZÜLÉKE ÉS MŰKÖDÉSI ELVE
Sugarak útja szabványos mikroszkóp fényforrás kondenzátor minta lencse szemlencse szem Sugarak útja modern mikroszkópban fényforrás mintagyűjtő kondenzátor szemlencse p képe a mintáról. A MIKROSZKÓP KÉSZÜLÉKE ÉS MŰKÖDÉSI ELVE
 Lencse nyitási szöge: MIKROSZKÓP FELBONTÁSA Rayleigh-képlet: A mikroszkóp térfelbontása a minimális távolság a kép két pontja között, amelyet akkor alkot, amikor azok külön-külön láthatók. ahol a használt fény hullámhossza, n a közeg törésmutatója és a lencse nyitási szöge. fényforrás mintagyűjtő kondenzátor lencse szemlencse p mintakép Abbe képlete: ahol NA a lencse numerikus apertúrája, egyenlő n sin (/2). NAd 61, 0 2/sin 2 n d
Lencse nyitási szöge: MIKROSZKÓP FELBONTÁSA Rayleigh-képlet: A mikroszkóp térfelbontása a minimális távolság a kép két pontja között, amelyet akkor alkot, amikor azok külön-külön láthatók. ahol a használt fény hullámhossza, n a közeg törésmutatója és a lencse nyitási szöge. fényforrás mintagyűjtő kondenzátor lencse szemlencse p mintakép Abbe képlete: ahol NA a lencse numerikus apertúrája, egyenlő n sin (/2). NAd 61, 0 2/sin 2 n d
 2 114 n. NA ndz Mikroszkóp mélységfelbontás – fókuszmélység. Young képlete:
2 114 n. NA ndz Mikroszkóp mélységfelbontás – fókuszmélység. Young képlete:
 0,2 mm átmérőjű lyukon áthaladó 650 nm hullámhosszú lézersugár diffrakciója MIKROSZKÓP, MINT DIFRAKCIÓ ÁTALAKÍTÓ dxxuixuxfu. F)]2 bűn()2)2 bűn()2))
0,2 mm átmérőjű lyukon áthaladó 650 nm hullámhosszú lézersugár diffrakciója MIKROSZKÓP, MINT DIFRAKCIÓ ÁTALAKÍTÓ dxxuixuxfu. F)]2 bűn()2)2 bűn()2))