Опорно-двигательная система. Двигательные функции
Психофизиология двигательной активности
Двигательная активность человека имеет очень широкий диапазон - от мышечных координации, требуемых для грубой ручной работы или перемещения всего тела в пространстве, до тонких движений пальцев при операциях, выполняемых под микроскопом. Обеспечение всех видов двигательной активности осуществляется на основе движения двух потоков информации. Один поток берет начало на периферии: в чувствительных элементах (рецепторах), которые находятся в мышцах, суставных сумках, сухожильных органах. Через задние рога спинного мозга эти сигналы поступают вверх по спинному мозгу и далее в разные отделы головного мозга.
С семидесятых годов были разработаны различные методы активного облегчения передвижения с использованием реальных объектов и обучения различным возможностям охвата и схватывания с упражнениями прогрессивной сложности, которые связаны с различными формами манипуляций. Рода Эрхардт, профессиональный терапевт, подробно рассказал о разработке последовательности для выпуска объектов.
Было опубликовано множество исследований с многообещающими результатами для улучшения функции верхних конечностей с помощью рестрикционной терапии здоровой стороны. Тем не менее, недавний систематический обзор у детей с гемиплегией предостерегает от его широкого использования, что указывает на большее количество исследований с хорошим методологическим дизайном. Было показано, что двусторонняя или бимануальная тренировка у пациентов с гемиплегией улучшает координацию между обеими руками, а также изолированно на пострадавшей стороне.
Взятые в совокупности сигналы от перечисленных образований образуют особый вид чувствительности - проприорецепцию. Хотя в сознании человека эта информация не отражается, благодаря ей мозг в каждый текущий момент времени имеет полное представление о том, в каком состоянии находятся все его многочисленные мышцы и суставы. Эта информация формируют схему или образ тела. Не имея такого интегрального образования, человек не мог бы планировать и осуществлять ни одно движение. Схема тела - исходное основание для реализации любой двигательной программы. Ее планирование, построение и исполнение связано с деятельностью двигательной системы.
Эти функциональные применения относятся к активации парализованных мышц в точной последовательности и величине для непосредственного выполнения функциональных задач. Отсюда и концепция нейропротезирования, применение которой позволяет заменить двигательную функцию верхних и нижних конечностей для задач самопомощи и подвижности, функции мочевого пузыря и дыхательного контроля.
Целенаправленная биологическая обратная терапия широко эффективна. Технология, называемая виртуальной реальностью, предлагает реальный опыт, улучшая результаты классических методов био-обратной связи. Однако истинные терапевтические преимущества этих систем еще не доказаны хорошо разработанными клиническими испытаниями.
В двигательной системе основной поток информации направлен от двигательной зоны коры больших полушарий - главного центра произвольного управления движениями - к периферии, т. е. к мышцам и другим органам опорно-двигательного аппарата, которые и осуществляют движение.
Строение двигательной системы
Существуют два основных вида двигательных функций: поддержание положения (позы) и собственно движение. В реальности разделить их достаточно сложно. Движения без одновременного удержания позы столь же невозможны, как удержание позы без движения.
Эти устройства известны как интерфейсы компьютерного мозга. Существуют исследования, которые пытаются улучшить знания о корковой физиологии, которая поддерживает человеческое намерение и обеспечивает сигналы для более сложного контроля, происходящего от церебрального сигнала.
Теории моторного обучения должны быть основой для восстановления двигателя. Исследования с хорошим методологическим дизайном продемонстрировали, что перенос терапевтических вех, достигнутых у неврологического пациента, в контексты, связанные с предметом, добавление изменчивости, активное участие, предоставление пациенту возможности совершать ошибки, давать обратную связь и стимулировать мотивацию, поскольку аспекты фундаментальным в сокращении функционального дефицита у неврологического пациента.
Нервные структуры, отвечающие за регуляцию положения тела в пространстве и движения, находятся в разных отделах ЦНС - от спинного мозга до коры больших полушарий. В их расположении прослеживается четкая иерархия, отражающая постепенное совершенствование двигательных функций в процессе эволюции.
У большинства животных в процессе эволюции выработались сложные локомоторные системы, позволяющие им достаточно успешно добывать пищу, спасаться от хищников, находить брачных партнеров и осваивать новые территории.
Таким образом, исследовательские линии, используя эти концепции, могут быть интересны при использовании новых методов и технологий в нейрореабилитации. Институт промышленной автоматизации Высшего совета научных исследований. Авторы не объявляют конфликта интересов.
Введение в нервную систему. Нервная система является правителем и координатором всех сознательных и бессознательных функций организма. Он состоит из цереброспинальной системы, нервов и вегетативной или автономной системы. Мозг - главная область интеграции нервной системы; это место, где хранятся воспоминания, мысли задуманы, генерируются эмоции и другие функции, связанные с нашей психикой и сложным контролем нашего тела. Чтобы выполнить эти сложные действия, сам мозг делится на многие функциональные части.
Локомоция стала возможной в результате формирования, взаимодействия и координированной работы нервной, мышечной и скелетной систем. Мышцы, участвующие в локомоции, работают как устройства, преобразующие химическую энергию в механическую. Мышцы способны сокращаться и при этом приводить в движение системы рычагов, составляющие часть скелета. Благодаря координированной работе рычагов, животное перемещается. Скелетно-мышечная система обеспечивает также поддержание позы и находится под общим контролем ЦНС.
Центральная нервная система выполняет самые высокие функции, поскольку она служит и удовлетворяет жизненные потребности и дает ответ на стимулы. Он выполняет три основных действия: выявление стимулов, передачу информации и общую координацию. Используя около 20% общего объема кровоснабжения тела, мозг является ключевым органом всего этого процесса. Их различные структуры регулируют чувствительность, движения, интеллект и функционирование органов. Его внешний слой, кору головного мозга, обрабатывает полученную информацию, сравнивает ее с хранимой информацией и преобразует ее в полезный, реальный и сознательный материал.
Самый низший уровень в организации движения связан с двигательными системами спинного мозга. В спинном мозге между чувствительными нейронами и мотонейронами, которые прямо управляют мышцами, располагаются вставочные нейроны, образующие множество контактов с другими нервными клетками. От возбуждения вставочных нейронов зависит, будет ли то или иное движение облегчено или заторможено. Нейронные цепи или рефлекторные дуги, лежащие в основе спинальных рефлексов, - это анатомические образования, обеспечивающие простейшие двигательные функции. Однако их деятельность в значительной степени зависит от регулирующих влияний выше расположенных центров.
Он обычно упоминается в разговорной речи как «мозг» для обозначения цефала теленима. Это включает весь мозг за вычетом промежуточного мозга и ствола мозга. Таким образом, теленцефалон образован корой головного мозга, базальными ганглиями и лимбической системой.
Периферическая нервная система представляет собой разветвленную сеть нервов. Его волокна двух типов: афферентные волокна для передачи чувствительной информации в спинной мозг и мозг и эфферентные волокна для передачи сигналов двигателя от центральной нервной системы к периферии, особенно поперечно-полосатых мышц. Некоторые периферические нервы имеют прямое происхождение в базальной области самого мозга и в основном иннервируют голову; Вместе они называются черепными нервами. Остальные периферические нервы - это спинномозговые нервы, каждый из которых распространяется на одну сторону спинного мозга через межпозвонковое отверстие в каждом позвоночнике.
Высшие двигательные центры находятся в головном мозге и обеспечивают построение и регуляцию движений. Двигательные акты, направленные на поддержание позы, и их координация с целенаправленными движениями осуществляется в основном структурами ствола мозга, в то же время сами целенаправленные движения требуют участия высших нервных центров. Побуждение к действию, связанное с возбуждением подкорковых мотивационных центров и ассоциативных зон коры, формирует программу действия. Образование этой программы осуществляется с участием базальных ганглиев и мозжечка, действующих на двигательную кору через ядра таламуса. Причем мозжечок играет первостепенную роль в регуляции позы и движений, а базальные ганглии представляют собой связующее звено между ассоциативными и двигательными областями коры больших полушарий (рис. 11.1).
Когда сигнал передается на нервном волокне, мембранный потенциал претерпевает ряд изменений, называемых потенциалом действия. Импульс распространяется по всему нервному волокну, и через эти импульсы нервное волокно передает информацию от одной части тела к другой.
Из нейронов, характерных для мозга или спинного мозга, основными его частями являются. Из этого растут другие части нейрона. Кроме того, тело клетки обеспечивает большую часть питания, которое требуется для сохранения жизни всего нейрона. Это много разветвлений клеточного тела. Большинство сигналов, подлежащих передаче, вводятся дендритами. Дендриты каждого нейрона обычно получают сигналы от тысяч точек соприкосновения с другими нейронами, которые называются синапсами.
Рис.11.1 Общий план организации двигательной системы.
Важнейшие двигательные структуры и их основные взаимосвязи указаны в левом столбце. Для простоты все чувствительные пути объединены вместе (кружок слева). В среднем столбце перечислены самые главные и твердо установленные функции, обнаруженные при раздельном изучении каждой из этих структур. В правом столбце указано, каким образом эти функции связаны с возникновением и выполнением движения. Следует обратить внимание на то, что базальные ганглии и мозжечок расположены на одном уровне, а двигательная кора участвует в превращении программы движения в его осуществление (по Дж. Дуделу с соавт., 1985)
Это часть нейрона, называемого нервным волокном. Аксоны передают нервные сигналы в следующую нервную клетку шейки матки или спинного мозга или в мышцы и железы в самых периферических частях тела. Концы аксонов и синапсы. В конце каждой из этих ветвей есть специализированный аксон-терминал, который в центральной нервной системе называется синаптической кнопкой из-за ее появления. Эта точка контакта между кнопкой и мембраной называется синапсом.
В центре этого волокна находится аксон, который передает нервный импульс. Вокруг аксона находится оболочка Шванна. Эта оболочка осаждается клетками Шванна, которые находятся вдоль периферических нервов, и обеспечивает электрическую изоляцию к аксонам.
Моторная или двигательная кора расположена непосредственно кпереди от центральной борозды. В этой зоне мышцы тела представлены топографически, т. е. каждой мышце соответствует свой участок области. Причем мышцы левой половины тела представлены в правом полушарии, и наоборот.
Двигательные пути, идущие от головного мозга к спинному, делятся на две системы: пирамидную и экстрапирамидную. Начинаясь в моторной и сенсомоторной зонах коры больших полушарий, большая часть волокон пирамидного тракта направляется прямо к эфферентным нейронам в передних рогах спинного мозга. Экстрапирамидный тракт, также идущий к передним рогам спинного мозга, передает им эфферентную импульсацию, обработанную в комплексе подкорковых структур (базальных ганглиях, таламусе, мозжечке).
Поскольку эта мембрана содержит большое количество жильного вещества миелина, изолирующая мембрана вокруг аксона называется «миелиновая оболочка». Милин обеспечивает отличную электрическую изоляцию для аксона. За первой ячейкой Шванна второй конверт обернут вокруг аксона. Соединение между двумя ячейками Шванна называется узелом Ранвье. В этом узле имеется тонкое пространство внеклеточной жидкости между двумя шванновскими клетками, и через эти пространства могут проходить небольшие количества ионов.
Поэтому узелка Ранвье очень важна для передачи нервных импульсов миелиновыми нервными волокнами. Импульсы передаются вдоль миелинового нерва с помощью процесса, называемого соляционной проводимостью, который деполяризует первый узел Ранвье. Это приводит к тому, что электрический ток направляется на следующий узел Ранвиера. Импульс «перескакивает» с одного узла на другой, который является процессом, называемым соляционной проводимостью.
Классификация движений
Все многообразие форм движения животных и человека основывается на физических законах перемещения тел в пространстве. При классификации движений необходимо учитывать конкретные целевые функции, которые должна выполнять двигательная система. В самом общем виде таких функций четыре: 1) поддержание определенной позы; 2) ориентация на источник внешнего сигнала для его наилучшего восприятия; 3) перемещения тела в пространстве; 4) манипулирование внешними вещами или другими телами. Иерархия уровней мозгового управления движениями также находится в зависимости от требований к структуре движения. Установлено, что подкорковый уровень связан с набором врожденных или автоматизированных программ.
Соляционное вождение ценно по двум причинам. Скорость проводимости на волокне увеличивается многократно. Миелиновая оболочка значительно уменьшает количество энергии, требуемой нервом для передачи импульса. Нервная ткань головного мозга, спинного мозга или периферических нервов содержит два основных типа клеток.
Нейроны, которые проводят сигналы в нервной системе, и из которых около 000 миллионов общесистемных. Поддерживают клетки и изоляцию, которые удерживают нейроны на месте и предотвращают распространение сигналов между этими клетками и их межклеточными структурами, которые в совокупности называются нейроглиями. В периферической нервной системе клетки с этой функцией называются клетками Шванна.
Автоматизированные и произвольные движения. Проблема разделения указанных категорий движения сложна. Во многих случаях грань между автоматизированным и произвольно контролируемым действием очень подвижна. Обучение устойчивым двигательным навыкам представляет собой переход от постоянно контролируемой цепочки более или менее осознанно выполняемых двигательных действий к автоматизированной слитной «кинетической мелодии», которая исполняется со значительно меньшими энергетическими затратами. В то же время достаточно небольшого изменения хотя бы одного из звеньев «кинетической мелодии», чтобы она перестала быть полностью автоматизированной, и для ее новой автоматизации требуется вновь вмешательство произвольной регуляции.
В организме нервные волокна обычно стимулируются физическими и химическими средствами. Например, давление на определенные нервные окончания кожи, механически растягивает эти окончания. Жара и простуда, повреждение тканей, такие как резка кожи и чрезмерное растяжение ткани, могут создавать болезненные импульсы.
Импульсы в центральной нервной системе передаются от одного нейрона к другому главным образом химическими средствами. Нервные окончания первого нейрона выделяют химическое вещество, называемое передатчиком, которое, в свою очередь, возбуждает второй нейрон.
Для того чтобы избежать трудностей, возникающих при попытках разделить двигательные акты на «автоматические» и «волевые», английский невропатолог Х. Джексон в начале века предложил иерархическую классификацию всех двигательных актов (т. е. движений и их комплексов) от «полностью автоматических» до «совершенно произвольных». Эта классификация оказывается полезной и в настоящее время. Так, например, дыхание представляет собой в значительной степени автоматический комплекс движений грудной клетки, мышц плечевого пояса и диафрагмы, сохраняющийся даже при самом глубоком сне и в состоянии наркоза, когда все остальные движения полностью подавлены. В случае, если при помощи тех же самых мышц осуществляется кашель или небольшие движения туловища, то подобный двигательный акт «менее автоматичен». В то же время при пении или речи эти мышцы участвуют уже в «совершенно неавтоматическом» движении. Из данного примера ясно также, что «более автоматические» движения связаны главным образом с врожденными центральными поведенческими программами, тогда как «менее автоматические» или «совершенно произвольные» движения появляются в процессе накопления жизненного опыта.
Большие миелиновые волокна очень быстро передают нервные сигналы. Эти сигналы регулируют быструю мышечную активность или передают чувствительные сигналы, очень важные для мозга. С другой стороны, амиленовые волокна контролируют структуры, такие как кровеносные сосуды, а также передают большое количество некритической сенсорной информации в мозг, такие как признаки грубого прикосновения со всех областей кожи, сигналы давления с поверхности тела, или болевые сигналы непрерывного типа из любого сайта организма.
Синапс представляет собой соединение между двумя нейронами. Через это соединение сигналы передаются от одного нейрона к другому. Синапс обладает способностью передавать некоторые сигналы и отвергать других, и поэтому является ценным сайтом центральной нервной системы, чтобы выбрать, что произойдет. Из-за этой передачи переменного сигнала синапс может быть единственным важнейшим детерминантом функционирования центральной нервной системы.
Ориентационные движения. Система движений такого рода, связана, во-первых, с ориентацией тела в пространстве, и во-вторых, с установкой органов чувств в положение, обеспечивающее наилучшее восприятие внешнего стимула. Примером первого может служить функция поддержания равновесия, второго - движения фиксации взора. Фиксация взора выполняется в основном глазодвигательной системой. Изображение неподвижного или движущегося предмета фиксируется в наиболее чувствительном поле сетчатки. Координация движения глаз и головы регулируется специальной системой рефлексов.
Синапсы состоят из союзов между синаптическими кнопками и дендритами или сомой. Малые волокна - это много ветвей аксонов других нейронов. На уровне синапса некоторые синаптические кнопки выделяют захватывающее передающее вещество, а другие выделяют тормозное передающее вещество; поэтому некоторые из этих терминалов возбуждают нейрон, а другие ингибируют его.
Синаптическая кнопка рядом с соматической мембраной нейрона. Это прекращение имеет много небольших везикул, содержащих вещество передатчика, и когда импульс нерва поступает на синаптическую кнопку, мгновенно изменяется структура мембранной кнопки, позволяя некоторым из этих пузырьков разряжать передающее вещество в синаптический канал, узкое пространство между кнопки и нейронной мембраны.
Управление позой. Поза тела определяется совокупностью значений углов, образуемых суставами тела животного или человека в результате ориентации в поле тяготения. Механизм позы складывается из двух составляющих: фиксации определенных положений тела и конечностей и ориентации частей тела относительно внешних координат (поддержание равновесия). Исходная поза тела накладывает некоторые ограничения на последующее движение. К низшим механизмам управления позой относятся спинальные, шейные установочные и некоторые другие рефлексы, к высшим - механизмы формирования «схемы тела».
Термином «схема тела» обозначают систему обобщенной чувствительности пространственных координат и взаимоотношений отдельных частей тела в покое и при движении. Общую «карту» тела для каждого полушария мозга обычно представляют в виде «гомункулуса». Топографически распределенная по поверхности коры чувствительность всего тела составляет ту основу, из которой путем объединения формируются целостные функциональные блоки крупных отделов тела. Эти интегративные процессы завершаются у взрослого организма и представляют собой закодированное описание взаиморасположения частей тела, которые используются при выполнении автоматизированных стереотипных движений.
Базой этих процессов служит анатомически закрепленная «карта» тела, поэтому такие процессы составляют лишь основу статического образа тела. Для его формирования необходимо соотнести эту информацию с положением тела по отношению к силе земного притяжения и взаиморасположением функциональных блоков тела в системе трех пространственных плоскостей. Вестибулярная система воспринимает перемещение всего тела вперед - назад, вправо - влево, вверх - вниз, а соответствующая информация поступает в теменные зоны коры, где происходит ее объединение с информацией от скелетно-мышечного аппарата и кожи. Туда же поступает импульсация от внутренних органов, которая также участвует в создании на бессознательном уровне особого психофизиологического образования - статического образа тела.
Таким образом, статический образ тела представляет собой систему внутримозговых связей, основанную на врожденных механизмах и усовершенствованную и уточненную в онтогенезе. Выполняя ту или иную деятельность, человек меняет взаиморасположение частей тела, а, обучаясь новым двигательным навыкам, он формирует новые пространственные модели тела, которые и составляют основу динамического образа тела. В отличие от статического, динамический образ тела имеет значение лишь для данного конкретного момента времени и определенной ситуации, при изменении которой он сменяется новым образом тела. Динамический образ базируется на текущей импульсации от чувствительных элементов кожи, мышц, суставов и вестибулярного аппарата. Не исключено, что скорость и точность формирования динамического образа тела - фактор, определяющий способность человека быстро овладевать новыми двигательными навыками.
В мозге происходит постоянное взаимодействие того и другого образов тела, осуществляется сличение динамического образа с его статическим аналогом. В результате этого формируется субъективное ощущение позы, отражающее не только положение тела в данный момент времени, но и возможные его изменения в непосредственном будущем. Если согласование не достигнуто, то вступают в действие активные механизмы перестройки позы. Итак, для того чтобы сменить позу, необходимо сравнить закодированный в памяти статический образ тела с его конкретной вариацией - динамическим образом тела.
Управление локомоцией. Термин локомоция означает перемещение тела в пространстве из одного положения в другое, для чего необходима определенная затрата энергии. Развиваемые при этом усилия должны преодолеть прежде всего силу тяжести, сопротивление окружающей среды и силы инерции самого тела. На локомоцию влияют характер и рельеф местности. Во время локомоции организму необходимо постоянно поддерживать равновесие.
Типичные примеры локомоции - ходьба или бег, которые отличаются стереотипными движениями конечностей, причем для каждой формы локомоции характерны две фазы шага: фаза опоры и фаза переноса. Ходьба человека характеризуется походкой, т. е. присущим ему особенностями перемещения по поверхности. Походка оценивается по способу распределения по времени циклических движений конечностей, длительностью опорной фазы и последовательностью перемещения опорных конечностей.
В спинном мозге обнаружена цепь нейронов, выполняющая функции генератора шагания. Она ответственна за чередование периодов возбуждения и торможения различных мотонейронов и может работать в автоматическом режиме. Элементарной единицей такого центрального генератора является генератор для одной конечности. Не исключено, что у каждой мышцы, управляющей одним суставом, есть собственный генератор. Когда человек движется, такие генераторы работают в едином режиме, оказывая друг на друга возбуждающее влияние.
Как известно, спинной мозг находится под непрерывным контролем высших двигательных центров. По отношению к локомоции этот контроль преследует ряд целей: 1) быстро запускает локомоцию, поддерживает постоянную скорость или изменяет ее, если требуется, а также прекращает ее в нужный момент времени; 2) точно соразмеряет движение (и даже отдельный шаг) с условиями среды; 3) обеспечивает достаточно гибкую позу, чтобы соответствовать различным условиям передвижения, таким, например, как ползание, плавание, бег по снегу, перенос груза и т. д.
Очень важную роль в этом контроле играет мозжечок, который обеспечивает коррекцию и точность постановки конечностей на основе сравнения информации о работе спинального генератора и реальных параметров движений. Предполагается, что мозжечок программирует каждый следующий шаг на основе информации о предыдущем. Другой важнейший уровень мозга, куда направляется информация о характере выполнения движения, это большие полушария с их таламическими ядрами, стриопаллидарной системой и соответствующими зонами коры головного мозга.
Обратная связь. Большое значение на этих уровнях контроля локомоции имеет обратная связь, т. е. информация о результатах выполняемого движения. Она поступает от двигательных аппаратов к соответствующим мозговым центрам. Многие движения постоянно корректируются, благодаря показаниям соответствующих сенсорных датчиков, расположенных в скелетных мышцах и передающих информацию в разные отделы мозга вплоть до коры. Движения, базирующиеся на врожденных координациях, в меньшей степени требуют обратной связи от локомоторного аппарата. Наряду с этим все новые формы движения, в основе которых лежит формирование новых координационных отношений, всецело зависят от обратной связи со стороны двигательного аппарата.
Функциональная структура произвольного движения. Из вышеизложенного следует, что в обеспечении любого движения принимают участие разные компоненты, поэтому один из главных вопросов состоит в том, каким образом обеспечивается единовременность команды, поступающей к исполнительным аппаратам. Независимо от стратегии и тактики конкретного движения, основная задача системы, обеспечивающей программу, заключается в координации всех компонетов команды.
ЦНС
располагает некоторым числом генетически закрепленных программ (например, локомоторная программа шагания, базирующаяся на активности Спинальный
- относящийся к позвоночнику.");" onmouseout="nd();" href="javascript:void(0);">спинального
генератора). Такие простые программы объединяются в более сложные системы типа поддержания вертикальной позы. Подобное объединение происходит в результате обучения, которое обеспечивается благодаря участию передних отделов коры больших полушарий.
Самой сложной и филогенетически самой молодой является способность формировать последовательность движений и предвидеть ее реализацию. Решение этой задачи связано с фронтальной ассоциативной системой, которая запоминает и хранит в памяти такие последовательности движений. Высшим отражением этого кодирования у человека является вербализация, или словесное сопровождение, основных понятий движения.
Всеобщей закономерностью работы системы управления движениями является использование обратной связи. Сюда входит не только проприоцептивная обратная связь от начавшегося движения, но и Активация
- возбуждение или усиление активности, переход из состояния покоя в деятельное состояние.");" onmouseout="nd();" href="javascript:void(0);">активация
систем поощрения или наказания. Кроме того, включается и внутренняя обратная связь, т.е. информация об активности нижележащих уровней двигательной системы, или эфферентная копия самой двигательной команды. Этот вид обратной связи необходим для выработки новых двигательных координаций. Для движений различной сложности и скорости обратная связь может замыкаться на разных уровнях. Поэтому оба типа управления — программирование и слежение — могут сосуществовать в системе управления одним и тем же движением.
В заключение целесообразно привести высказывание выдающегося физиолога Н.А. Бернштейна о том, что движения "...ведет не пространственный, а смысловой образ и двигательные компоненты цепей уровня действий диктуются и подбираются по смысловой сущности предмета и того, что должно быть проделано с ним".
10.4. Электрофизиологические корреляты организации движения
Электрофизиологические методы используются для изучения разных сторон двигательной активности, и в первую очередь тех из них, которые недоступны прямому наблюдению. Ценную информацию о физиологических механизмах организации движения дают методы оценки взаимодействия зон коры мозга, анализ локальной ЭЭГ и потенциалов, связанных с движением, а также регистрация активности нейронов.
Исследование межзональных связей биопотенциалов мозга позволяет проследить динамику взаимодействия отдельных зон коры на разных этапах выполнения движения, при обучении новым двигательным навыкам, выявить специфику межзонального взаимодействия при разных типах движений.
Пространственная синхронизация (ПС)
, т.е. синхронная динамика электрических колебаний, регистрируемых из разных точек коры больших полушарий, отражает такое состояние структур мозга, при котором облегчается распространение возбуждения и создаются условия для межзонального взаимодействия. Метод регистрации ПС был разработан выдающимся отечественным физиологом М.Н. Ливановым.
Исследования ритмических составляющих ЭЭГ отдельных зон и их пространственно-временных отношений у человека во время выполнения произвольных движений дал реальную возможность подойти к анализу центральных механизмов функциональных взаимодействий, складывающихся на системном уровне при двигательной деятельности. Корреляционный анализ ЭЭГ, зарегистрированной во время выполнения ритмических движений, показал, что у человека в корковой организации движений принимают участие не только центры моторной коры, но также лобные и нижнетеменные зоны.
Обучение произвольным движениям и их тренировка вызывают перераспределение межцентральных корреляций корковых биопотенциалов. В начале обучения общее число центров, вовлеченных в совместную деятельность, резко возрастает, и усиливаются синхронные отношения ритмических составляющих ЭЭГ моторных зон с передними и задними Ассоциативные зоны коры
- зоны, которые получают информацию от рецепторов, воспринимающих раздражение различной модальности, и от всех проекционных зон.");" onmouseout="nd();" href="javascript:void(0);">ассоциативными
областями. По мере овладения движением общий уровень ПС значительно снижается, и, напротив, усиливаются связи моторных зон с нижнетеменными.
Важно отметить, что в процессе обучения происходит перестройка ритмического состава биопотенциалов разных зон коры: в ЭЭГ начинают регистрироваться медленные ритмы, совпадающие по частоте с ритмом выполнения движений. Эти ритмы в ЭЭГ человека получили название "меченых". Такие же меченые колебания были обнаружены у детей дошкольного возраста при совершении ими ритмических движений на эргографе.
Систематические исследования ЭЭГ человека во время осуществления циклической (периодически повторяющейся) и ациклической двигательной активности позволили обнаружить значительные изменения в динамике электрической активности коры больших полушарий. В ЭЭГ происходит усиление как локальной, так и дистантной синхронизации биопотенциалов, что выражается в нарастании мощности периодических составляющих, в изменениях частотного спектра авто- и кросскоррелограмм, в определенной сонастройке максимумов частотных спектров и функций Когерентность
- степень синхронизации частотных показателей ЭЭГ между различными отделами коры головного мозга.");" onmouseout="nd();" href="javascript:void(0);">когерентности
на одной и той же частоте.
ПС и время реакции.
Время реакции — один из наиболее простых двигательных показателей. Поэтому особый интерес представляет тот факт, что даже простая двигательная реакция может иметь различающиеся физиологические корреляты в зависимости от увеличения или сокращения ее длительности. Так, при сопоставлении картины межцентральных корреляционных отношений спектральных составляющих ЭЭГ мозга со временем простой двигательной реакции выяснилось, что перестройка пространственно-временных отношений ЭЭГ ассоциативных зон связана с временем реакции на заданный стимул. При быстрых реакциях у здорового человека чаще всего высокие корреляционные связи биопотенциалов возникали в обеих нижнетеменных областях (несколько больше с левым полушарием мозга). Если время реакции возрастало, это сопровождалось синхронизацией биопотенциалов в лобных отделах коры и из взаимодействия исключалась нижнетеменная область левого полушария. Кроме того, была обнаружена зависимость между величинами фазовых сдвигов Альфа-ритм
- основной ритм электроэнцефалограммы в состоянии относительного покоя, с частотой в пределах 8 - 14 Гц и средней амплитудой в 30 - 70 мкВ.");" onmouseout="nd();" href="javascript:void(0);">альфа-ритма
, зарегистрированного в лобных, прецентральных и затылочных областях мозга и скоростью простой двигательной реакции.
Важно отметить, что усиление синхронизации биопотенциалов наступает у человека уже в предрабочем состоянии в процессе сосредоточения перед двигательным действием, а также при мысленном выполнении движений.
ПС и специфика движения.
Кроме неспецифического усиления ПС биопотенциалов было отмечено ее выраженное избирательное нарастание между зонами коры, непосредственно участвующими в организации конкретного двигательного акта. Например, наибольшее сходство в электрической активности устанавливается: при движении рук — между лобной областью и моторным представительством мышц верхних конечностей; при движении ног — между лобной областью и моторным представительством мышц нижних конечностей. При точностных действиях, требующих тонкой пространственной ориентации и зрительного контроля (стрельба, фехтование, баскетбол), усиливаются взаимодействия между зрительными и моторными областями.
Была выявлена сложная динамика ПС биопотенциалов различных участков мозга у спортсменов при выполнении различных упражнений и показана зависимость нарастания взаимодействия ритмических составляющих ЭЭГ от режима двигательной деятельности, от квалификации спортсменов, от способности человека решать тактические задачи, от сложности ситуации. Так, у спортсменов высокой квалификации межцентральные взаимодействия выражены гораздо интенсивнее и локализованы более четко. Выяснилось также, что более сложные двигательные задачи требуют для своего успешного решения более высокого уровня пространственной синхронизации ритмов ЭЭГ, а время решения тактических задач коррелирует со скоростью нарастания межцентральных взаимодействий. При этом двигательный ответ следует после достижения максимума синхронности биопотенциалов в коре головного мозга.
В совокупности исследования ПС биопотенциалов мозга у человека позволили установить, что при выполнении простых и сложных двигательных актов во взаимодействия вступают разные центры мозга, образуя при этом сложные системы взаимосвязанных зон с фокусами активности не только в проекционных, но и в ассоциативных областях, особенно лобных и нижнетеменных. Эти межцентральные взаимодействия динамичны и изменяются во времени и пространстве по мере осуществления двигательного акта.
10.5. Комплекс потенциалов мозга, связанных с движениями
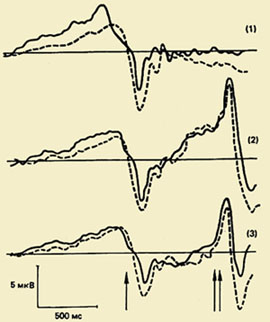 Одним из важных направлений в исследовании психофизиологии двигательного акта является изучение комплекса колебаний потенциалов мозга, связанных с движениями (ПМСД). Значение этого феномена для понимания физиологических механизмов организации движения очень велико, потому что изучение ПМСД позволяет выявить скрытую последовательность процессов, происходящих в коре мозга при подготовке и выполнении движения, и Хронометрия процессов переработки информации
- совокупность методов измерения длительности отдельных стадий в процессе переработки информации на основе измерения физиологических показателей, в частности латентных периодов компонентов вызванных и событийно-связанных потенциалов.");" onmouseout="nd();" href="javascript:void(0);">хронометрировать
эти процессы, т.е. установить временные границы их протекания.
Одним из важных направлений в исследовании психофизиологии двигательного акта является изучение комплекса колебаний потенциалов мозга, связанных с движениями (ПМСД). Значение этого феномена для понимания физиологических механизмов организации движения очень велико, потому что изучение ПМСД позволяет выявить скрытую последовательность процессов, происходящих в коре мозга при подготовке и выполнении движения, и Хронометрия процессов переработки информации
- совокупность методов измерения длительности отдельных стадий в процессе переработки информации на основе измерения физиологических показателей, в частности латентных периодов компонентов вызванных и событийно-связанных потенциалов.");" onmouseout="nd();" href="javascript:void(0);">хронометрировать
эти процессы, т.е. установить временные границы их протекания.
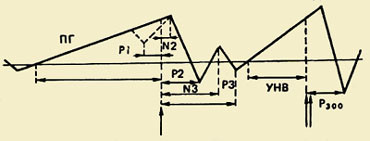
Компонентный состав ПМСД. Впервые этот комплекс, отражающий процессы подготовки, выполнения и оценки движения был зарегистрирован в 60-е годы. Оказалось, что движению предшествует медленное отрицательное колебание — потенциал готовности (ПГ). Он начинает развиваться за 1,5 - 0,5 с до начала движения. Этот компонент регистрируется преимущественно в центральных и лобно-центральных отведениях обоих полушарий. За 500-300 мс до начала движения ПГ становится асимметричным — его максимальная амплитуда наблюдается в прецентральной области, Контралатеральный - относящийся к противоположной стороне тела.");" onmouseout="nd();" href="javascript:void(0);">контралатеральной движению. Примерно у половины взрослых испытуемых на фоне этого медленного отрицательного колебания незадолго до начала движения регистрируется небольшой по амплитуде положительный компонент. Он получил название "премоторная позитивность" (ПМП). Следующее по порядку быстро нарастающее по амплитуде отрицательное колебание, так называемый моторный потенциал (МП), начинает развиваться за 150 мс до начала движения и достигает максимальной амплитуды над областью моторного представительства движущейся конечности в коре головного мозга. Завершается этот комплекс потенциалов положительным компонентом примерно через 200 мс после начала движения.
Функциональное значение компонентов.
Принято считать, что потенциал готовности (ПГ) возникает в моторной коре и связан с процессами планирования и подготовки движения. Он относится к классу медленных отрицательных колебаний потенциала мозга, возникновение которых объясняют активацией нейрональных элементов соответствующих участков коры.
Гипотезы относительно функционального значения ПМП различны.
Это колебание рассматривают и как отражение подачи центральной команды от коры к мышцам, и как результат релаксации коры после завершения определенного этапа организации движения, и как отражение процессов подавления ассоциированных движений другой конечности, и как обратную связь от мышечных афферентов. В настоящее время некоторые авторы полагают, что ПМП являются лишь отражением начала моторного потенциала.
При регистрации МП у обезьян в составе МП были выделены два субкомпонента. Первый субкомпонент соотносят с активацией моторной коры, связанной с инициацией движения (синаптическая активность пирамидных нейронов), а второй — с активацией полей 2.3 и 4 Бродмана поля
- отдельные участки коры больших полушарий мозга, отличающиеся по своему клеточному строению (цитоархитектонике) и функциям. Например, поля 17,18,19 - зрительные области коры больших полушарий, которые имеют разное строение и функции в обеспечении зрительного восприятия.");" onmouseout="nd();" href="javascript:void(0);">по Бродману
. Регистрация МП у человека, больного эпилепсией, позволила выделить в нем три компонента. Первый компонент был назван потенциалом инициации. Он имеет высокую амплитуду и возникает после начала движения в прецентральной контралатеральной коре. Второй, возникающий после начала миограммы, более локализованный в контралатеральном соматосенсорном поле может быть связан как с инициацией движения, так и с сенсорной обратной связью. Третий компонент отражает импульсацию, поступающую с мышечных афферентов в кору.
Следующий за МП позитивный потенциал рассматривается как отражение обратной афферентации, поступающей с периферических рецепторов, восходящей активности от моторных центров, операции сравнения между моторной программой и нейрональной картиной ее исполнения или процессов релаксации коры после выполнения движения.
Волна ожидания. Помимо ПМСД, описан еще один электрофизиологический феномен, который по своей сути близок потенциалу готовности. Речь идет об отрицательном колебании потенциала, регистрируемого в передних отделах коры мозга в период между действием предупреждающего и пускового (требующего реакции) сигналов. Это колебание имеет ряд названий: волна ожидания, Е-волна, условное негативное отклонение (УНВ). Е-волна возникает через 500 мс после предупредительного сигнала, ее длительность растет с увеличением интервала между первым и вторым стимулами. Амплитуда Е-волны растет прямо пропорционально скорости двигательной реакции на пусковой стимул. Она увеличивается при напряжении внимания и увеличении волевого усилия, что свидетельствует о связи этого электрофизиологического явления с механизмами произвольной регуляции двигательной активности и поведения в целом.
10.6. Нейронная активность
Функциональные кортикальные колонки.
В моторной зоне коры у человека имеются так называемые гигантские пирамидные Беца-клетки
- пирамидные клетки коры головного мозга.");" onmouseout="nd();" href="javascript:void(0);">клетки Беца
, которые организованы в обособленные колонки. Пирамидные клетки, выполняющие сходные функции, расположены рядом друг с другом, иначе было бы трудно объяснить точную соматотопическую организацию коры. Такие двигательные колонки способны возбуждать или тормозить группу функционально однородных Мотонейрон
(двигательный нейрон) - нервная клетка, аксон которой иннервирует мышечные волокна.");" onmouseout="nd();" href="javascript:void(0);">мотонейронов
.
Регистрация активности одиночных пирамидных клеток с помощью вживленных микроэлектродов у животных, выполняющих различные движения, позволила установить принципиально важный факт. Нейроны коры, регулирующие деятельность какой-либо мышцы, не сосредоточены в пределах только одной колонки. Двигательная колонка в значительной степени представляет собой функциональное объединение нейронов, регулирующих деятельность нескольких мышц, действующих на тот или иной сустав. Таким образом, в колонках пирамидных нейронов моторной коры представлены не столько мышцы, сколько движения.
Нейронные коды моторных программ.
Кодирование информации в нейроне осуществляется частотой его разрядов. Анализ импульсной активности нейронов при выработке у животных различных моторных программ показал, что в их построении участвуют нейроны разных отделов двигательной системы, выполняя при этом специфические функции. По некоторым представлениям включение моторных программ происходит благодаря активации так называемых командных нейронов. Командные нейроны находятся, в свою очередь, под контролем высших корковых центров. Торможение
- нервный процесс, противоположный возбуждению; проявляется в ослаблении или прекращении деятельности, специфической для данной системы организма.");" onmouseout="nd();" href="javascript:void(0);">Торможение
командного нейрона приводит к остановке контролируемой им программы, возбуждение, напротив, к активизации нервной цепи и актуализации моторной программы.
Вовлечение командных нейронов в целостную деятельность мозга определяется текущей мотивацией и конкретной двигательной программой направленной на удовлетворение этой мотивации. Двигательная программа, чтобы носить приспособительный характер, должна учитывать все сигнально значимые компоненты внешней среды, относительно которых совершается целенаправленное движение, т.е. строиться на принципе мультисенсорной Конвергенция
- объединение аксонов группы нейронов, возникающее благодаря образованию синапсов на одном и том же постсинаптическом нейроне.");" onmouseout="nd();" href="javascript:void(0);">конвергенции
.
Словарь терминов
- локомоция
- мотонейрон
- артикуляция
- активация
- когерентность
- Бродмана поля
- хронометрия процессов переработки информации
Вопросы для самопроверки
- Чем различаются статический и динамический образ тела?
- Какие структуры мозга играют решающую роль в организации произвольного движения?
- Как различаются функции пирамидной и экстрапирамидной систем?
- Что дают для понимания мозговых механизмов организации движения потенциалы мозга, связанные с движением?
Список литературы
- Батуев А.С. Функции двигательного анализатора. Л.: Наука, 1970.
- Батуев А.С. Высшая нервная деятельность. М.: Высшая школа, 1991.
- Бернштейн Н.А. Очерки по физиологии движений и физиологии активности. М.: Наука, 1966.
- Блум Ф., Лайзерсон А., Хофстедтер Л. Мозг, разум и поведение. М.: Мир, 1988.
- Дудел Дж., Рюэгг И., Шмидт Р., Яниг В. Физиология человека. Т. 1 / Под ред. Р. Шмидта и Г. Тевса. М.: Мир, 1985.
- Общий курс физиологии человека и животных / Под ред. А.А. Ноздрачева. М.: Высшая школа, 1991.
- Сологуб Е.Б. Электрическая активность мозга человека в процессе двигательной деятельности. Л.: Наука, 1973.
- Хризман Т.П. Движение ребенка и электрическая активность мозга. М.: Педагогика, 1973.
- Эвартс Э. Механизмы головного мозга, управляющие движением // Мозг. М.: Мир, 1982.
Темы курсовых работ и рефератов
- Учение Н.А. Бернштейна о строении движения.
- Психофизиологические типологии движений.
- Движения руки человека и их нейрофизиологические механизмы.
- Произвольные движения и их нейрофизиологические механизмы.
- Исторические этапы исследований потенциалов мозга, связанных с движениями.
- Роль нейронной активности в построении движений.



