Механизмы сна и бодрствования организма. Сон как особая активность мозга. Теории сна. Современные представления о природе сна. Значение сна. Нарушение цикла «сон-бодрствование. Реферат Высшая нервная деятельность: Сон и бодрствование Циркадианная теория с
Возбуждение и торможение, возникнув в определенном участке нервной системы, способны распространяться. Этот процесс, открытый еще в XIX веке, получил название иррадиации. Так, если раздражать электрическим током двигательную область коры, то постепенно в ответ вовлекается все большее количество мышц, а затем реакция выходит за пределы двигательной системы, например, повышается слюноотделение и учащается сердцебиение.
Зная механизмы работы мозга на клеточном уровне, легко понять, что в основе иррадиации лежит распространение ПД по аксонам возбуждающих либо тормозных нейронов. Учтем также процессы суммации, когда, повышая возбудимость синапсов, повторные нервные сигналы «прокладывают» себе путь, все более удаляясь от точки возникновения.
При снижении уровня возбуждения (торможения) наблюдается противоположный по отношению к иррадиации феномен - концентрация (сосредоточение) нервных процессов в исходном пункте. Важно, что при концентрации в определенном участке мозга нервный процесс одновременно вызывает (индуцирует) противоположный нервный процесс в окружающих зонах. В том случае, когда возбуждение индуцирует торможение, говорят об отрицательной индукции. В обратном случае, когда торможение индуцирует возбуждение, индукция положительная. Особенно важное биологическое значение имеет отрицательная индукция. Смысл ее таков: сильный очаг возбуждения способен тормозить другие аналогичные очаги в нервной системе (так реализуется принцип доминанты).
Наиболее широкие процессы иррадиации возбуждения и торможения в ЦНС обеспечивают системы сна и бодрствования.
У человека выявлено большое число физиологических показателей, которые изменяются с периодом в 24 ч (температура тела, артериальное давление, выделение гормонов), причем такая ритмичность сохраняется даже при неизменности всех внешних факторов (например, если человек долгое время находится в пещере). Это говорит о том, что в процессе эволюции в условиях планеты Земля, совершающей оборот вокруг своей оси за 24 ч, организмы приспособились к такому ритму и появились эндогенные (внутренние) механизмы, обусловливающие его сопровождение. Запускаемые ими ритмы принято называть околосуточными, или циркадианными (лат. circa - около и dies - день). Наиболее выражен суточный цикл сон - бодрствование. Он реализуется как за счет безусловно-рефлекторной основы, главные компоненты которой относятся к группе витальных врожденных реакций, так и за счет определенных адаптивных процессов, которые и будут рассмотрены ниже.
Состояние бодрствования характеризуется высокой нервной активностью. В это время происходит взаимодействие организма с внешней средой, формирование и реализация различных рефлекторных реакций. Бодрствующее состояние поддерживается особыми центрами бодрствования.
Сон - это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью.
Павлов предлагал выделять два вида сна - активный и пассивный. Первый он определял как возникающий при отсутствии (недостатке) внешних раздражителей. Соответственно для поддержания бодрствующего состояния часто необходим некоторый минимум сенсорных стимулов.
Активный сон, по Павлову, возникает как результат работы особых центров сна. Павлов представлял активный сон в виде тормозного процесса, иррадиирующего из определенного утомленного центра коры и захватывающего весь головной мозг. Дальнейшие исследования показали, что гораздо чаще источником сна являются другие, более глубоко расположенные структуры головного мозга.
Кратко охарактеризуем основные центры бодрствования и сна головного мозга (рис. 4.25).
Уже в самых ранних исследованиях механизмов сна четко намечаются две основные точки зрения на эту проблему. Первая – сон возникает в результате активного процесса, возбуждения определенных структур («центров сна»), которое вызывает общее снижение функций организма (активные теории сна). Вторая – это пассивные теории сна, или теории деафферентации, согласно которым сон наступает пассивно в результате прекращения действия каких-то факторов, необходимых для поддержания бодрствования. Различия между этими направлениями удачно определил Н. Клейтман, писавший, что «заснуть» и «не суметь остаться бодрствующим», – это не одно и то же, так как первое предполагает активное действие, а второе – пассивное устранение активного состояния.
Первыми экспериментальными исследованиями, свидетельствующими о существовании центра сна, явились работы В. Гесса. Показав, что слабое электрическое раздражение четко ограниченной области промежуточного мозга у подопытных кошек вызывало сон со всеми подготовительными фазами (потягивание кошки, умывание, принятие характерной позы), В. Гесс высказал предположение, что существует центр, возбуждение которого обеспечивает наступление естественного сна. В дальнейшем опыты В. Гесса были подтверждены многочисленными исследователями, которые вызывали наступление сна у подопытных животных при помощи электрического и химического раздражения гипоталамуса и прилежащих структур, и теория о центре сна получила значительное признание.
Однако против подобного локализационистского объяснения механизма возникновения сна решительно выступал И.П. Павлов. Он рассматривал сон как результат торможения коры больших полушарий; при этом его теория сна не исключала участия и подкорковых структур в возникновении сна.
С. Рэнсон пришел к выводу о том, что гипоталамус является центром «интеграции эмоционального выражения» и сон наступает в результате периодического снижения активности этого центра бодрствования.
Открытие Дж. Моруцци и X. Мэгуном в 1949 г. восходящего активирующего влияния неспецифической ретикулярной системы (ВРАС) значительно усилило позиции пассивных теорий сна. Поддержание бодрствующего состояния объяснялось теперь тоническим влиянием ВРАС. Дальнейшие исследования привели к открытию и других активирующих систем – диффузной и специфической таламической систем и активирующих структур заднего гипоталамуса (см. гл. 8).
Одна из попыток создания единой теории сна была предпринята П.К. Анохиным . Состояние сна он представлял как результат проявления целостной деятельности организма, строго координирующей корковые и подкорковые структуры в единую функциональную систему. В своей гипотезе П.К. Анохин исходил из того, что гипоталамические «центры сна» находятся под тоническим угнетающим влиянием со стороны коры больших полушарий. Именно поэтому при ослаблении этого влияния вследствие снижения рабочего тонуса корковых клеток («активный сон» по Павлову) гипоталамические структуры как бы «высвобождаются» и определяют всю ту сложную картину перераспределения вегетативных компонентов, которая характерна для состояния сна. При этом гипоталамические центры оказывают угнетающее влияние на восходящую активирующую систему, прекращая доступ в кору всего комплекса активирующих воздействий (и наступает «пассивный сон» по Павлову). Эти взаимодействия представляются циклическими, поэтому состояние сна может быть вызвано искусственно (или в результате патологического процесса) воздействием на любую часть этого цикла (рис. 13.1).
В настоящее время после открытия целого ряда активирующих и синхронизирующих структур мозга, а также многочисленных пептидов и нейротрансмиттеров (см. далее), участвующих в регуляции цикла «сон–бодрствование», эта схема наполняется новым содержанием.
В 1953 г. Е. Азеринский и Н. Клейтман открыли феномен «быстрого» сна, и тем самым – новую эру в изучении сна. Если раньше пассивные и активные теории регуляции сна рассматривали бодрствование как состояние, противоположное сну, а сон сам по себе считался единым феноменом, то теперь идея монолитного сна оказалась разрушенной и стали изучаться механизмы как медленного, так и быстрого сна. В итоге в настоящее время регуляторные процессы медленного сна связываются со структурами промежуточного мозга, а быстрого сна – главным образом со стволовыми структурами моста.
В 60–70 гг. М. Жуве, основываясь на обширных исследованиях с пересечениями и повреждениями мозга, а также фармакологических и нейроанатомических данных, предложил моноаминергическую теорию регуляции цикла «сон–бодрствование», согласно которой медленный и быстрый сон связаны с активностью различных групп моноаминергических нейронов – в регуляцию медленного сна включены серотонинергические нейроны комплекса шва, в то время как норадренергические нейроны ответственны за наступление быстрого сна. Впоследствии было показано участие различных нейротрансмиттеров в регуляции медленного и быстрого сна. В табл. 13.1 представлены эти данные .
Различие механизмов медленного и быстрого сна подтверждается также и в нейрогуморальных концепциях сна, основоположником которых является А. Пьерон. Еще в начале текущего столетия, на основании результатов своих экспериментов на собаках, у которых сон вызывался введением спинномозговой жидкости других собак, лишенных сна в течение нескольких суток, А. Пьерон предположил, что наступление сна связано с накоплением в организме определенных веществ (гипнотоксинов). Впоследствии «фактор сна» многочисленные исследователи выделяли из спинномозговой жидкости, крови и мочи различных животных, и с каждым годом увеличивался список обнаруженных в организме веществ, связанных со сном. В табл. 13.2 представлены все пептиды, изучавшиеся на предмет влияния на сон. Р. Друкер-Колин и Н. Мерчант-Нэнси , суммировав полученные данные, объясняют обилие этих веществ тем, что все они действуют через посредство какого-то еще неизвестного механизма, ответственного за наступление сна, а единственного фактора сна в понимании А. Пьерона реально не существует.
Ко всем перечисленным веществам нужно добавить мелатонин, который выделяется эпифизом только ночью и также играет важную роль в поддержании сна (о механизме действия различных групп веществ на сон см. обзор ).
Таким образом, результаты обширных нейрофизиологических, нейрохимических и нейрогуморальных исследований свидетельствуют не только о сложности и многообразии взаимодействия различных факторов в регуляции цикла «сон–бодрствование», но и о различии механизмов медленного и быстрого сна.
А. Сон - это особая активность мозга, при которой у человека выключаются сознание и механизмы поддержания естественной позы, снижена чувствительность анализаторов. Рекомендуе мая продолжительность сна взрослого человека 7-8 часов в сутки.
Для оценки глубины сна обычно используют электроэнцефалограмму (ЭЭГ). По особенностям ЭЭГ, исходя из общепринятых стандартных критериев, выделяют четыре или пять его стадий (рис. 13.8).
Перед пробуждением спящий человек обычно проходит через особую фазу сна, характеризующуюся десинхронизацией ЭЭГ и эпизодами быстрых движений глаз (БДГ). Их можно наблюдать со стороны через сомкнутые веки спящего или записывать методами электроокулографии. БДГ настолько характерны для данной фазы, что ее называют сном с БДГ - быстрым сном, другие фазы называют медленным (синхронизированным) сном. Остальная мускулатура в фазе быстрого сна, как и во время медленного сна, практически атонична, если не считать иногда возникающих судорожных сокращений мышц лица или пальцев.
Порог пробуждения в БДГ-фазе сна примерно такой же, как и во время глубокого сна, однако ЭЭГ сходна с записываемой при бодрствовании или переходе ко сну, поэтому БДГ-сон получил также название парадоксального, или десинхронизированного.
На протяжении ночи последовательность стадий сна повторяется в среднем три - пять раз. Как правило, максимальная его
глубина при каждом таком цикле убывает к утру. В норме БДГ-сон повторяется примерно через каждые 1,5 ч и длится в среднем 20 мин, причем с каждым разом все больше.
Б. Сновидения - возникающие во сне образные представления, воспринимаемые как реальная действительность. Сновидения, очевидно, возникают главным образом во время сна с БДГ.
Факторы, побуждающие сновидения. 1. Предшествующая сну деятельность (дети продолжают «играть» во сне, исследователь ставит эксперименты и т. д.). Например, известному физиологу О. Леви приснилась модель опыта, с помощью которого он открыл медиаторный механизм передачи влияний с симпатического и парасимпатического нервов на сердце. Д. И. Менделееву сновидение помогло создать свою знаменитую таблицу. 2. Раздражители, действующие на организм во время сна. Так, если приложить горячую грелку к ногам спящего человека, может присниться сон, что он идет по раскаленному песку. 3. Избыточная импульсация от переполненных или больных внутренних органов может вызывать кошмарные сновидения.
В. Значение сна.
Сон обеспечивает отдых организма. В экспериментах М. М. Манассеиной (1892) было показано, что лишенные сна взрослые собаки погибали на 12-21-й день. Лишение сна щенков приводило их к гибели через 4-6 дней. Депривация сна человека в течение 116 часов сопровождалась нарушением поведения, повышением раздражительности, психическими расстройствами. Более значительно меняется поведение человека при лишении его медленного сна, в результате чего возникает повышенная возбудимость.
Сон играет важную роль в процессах метаболизма. Полагают, что медленный сон способствует восстановлению внутренних органов, поскольку через гипоталамус либерины воздействуют на гипофиз, способствуя освобождению гормона роста (ГР), который участвует- в биосинтезе белков в периферических тканях. Напротив, парадоксальный сон восстанавливает пластические свойства нейронов головного мозга, усиливает процессы в клетках нейроглии, которые обеспечивают нейроны питательными веществами и кислородом.
3. Сон способствует переработке и запоминанию информа ции. Информация, предъявляемая во время сна, не запомина ется, если только на ЭЭГ во время или после этого не появляется а-ритм (т. е. если человек не просыпается). Из всех проявлений активности мозга во время сна запоминается лишь последнее сно видение. С другой стороны, сон облегчает закрепление изучен-
ново материала. Если какая-то информация заучивается непосредственно перед засыпанием, то спустя 8 ч она вспоминается лучше (утро вечера мудренее).
4. Биологическое значение сна связано с приспособлением к изменению освещенности (день-ночь). Организм способен заранее приспособиться к ожидаемому воздействию со стороны внешнего мира, активность всех систем снижается в определенные часы согласно режиму труда и отдыха. К моменту пробуждения и в начале бодрствования активность органов и систем возрастает и соответствует уровню поведенческих реакций.
Г. Механизмы бодрствования и сна.
Переход от бодрствования ко сну предполагает два возможных пути. Прежде всего, не исключено, что механизмы, поддерживающие бодрствующее состояние, постепенно «утомляются». В соответствии с такой точкой зрения, сон - это пассивное явление, следствие снижения уровня бодрствования. Однако не исключено и активное торможение обеспечивающих бодрствование механизмов. Важную роль в регуляции цикла сон - бодрствование играет ретикулярная формация ствола мозга, где находится множество диффузно расположенных нейронов, аксоны которых идут почти ко всем областям головного мозга, -за исключением неокортекса. Ее роль в цикле сон - бодрствование была исследована в конце 1940-х гг. Г. Моруцци и Н. Мэгуном. Они обнаружили, что высокочастотное электрическое раздражение этой структуры у спящих кошек приводит к их мгновенному пробуждению. И напротив, повреждения ретикулярной формации вызывают постоянный сон, напоминающий кому; перерезка же только сенсорных трактов, проходящих через ствол мозга, такого эффекта не дает.
Серотонинергические нейроны также играют весьма важную роль в регуляции бодрствования и сна. В верхних отделах ствола мозга есть две области - ядра шва и голубое пятно, у нейронов которых такие же обширные проекции, как и у нейронов ретикулярной формации, т. е. достигающие многих областей ЦНС. Медиатором в клетках ядер шва служит серотонин (5-гидрокситрипта-мин, 5-НТ), а голубого пятна - норадреналин. Разрушение ядер шва у кошки приводит к полной бессоннице в течение нескольких дней; за несколько следующих недель сон нормализуется. Частичная бессонница может быть также вызвана подавлением синтеза 5-НТ я-хлорфенилаланином. Ее можно устранить введением 5-гид-рокситриптофана, предшественника серотонина (последний не проникает через гематоэнцефалический барьер). Двустороннее разру-
шение голубого пятна приводит к полному исчезновению БДГ-фаз, не влияя на медленноволновой сон. Истощение запасов серотонина и норадреналина под влиянием резерпина вызывает, как и следовало ожидать, бессонницу.
Были сделаны попытки обнаружить особые вещества либо после длительного лишения сна, либо у спящего человека. Первый из этих подходов основан на предположении о том, что фактор(ы) сна во время бодрствования накапливаются до вызывающего сон уровня, а второй - на гипотезе, согласно которой они образуются или выделяются во сне.
Оба подхода дали определенные результаты. Так, при проверке первой гипотезы из мочи и спинномозговой жидкости человека и животных был выделен небольшой глюкопептид - фактор 5, вызывающий медленноволновой сон при введении другим животным.
Однако сросшиеся девочки-близнецы могли спать порознь, что свидетельствует о второстепенной роли гуморальных факторов и решающей роли нервной системы в развитии сна.

 Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей в изоляции от внешнего мира и использующего только искусственное освещение. Темные полосы - периоды сна, светлые - бодрствование.
Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей в изоляции от внешнего мира и использующего только искусственное освещение. Темные полосы - периоды сна, светлые - бодрствование.
 Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно получает вход от зрительной системы и отвечает за восприятие светлого времени суток как времени бодрствования, а также поддерживает суточный ритм.
Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно получает вход от зрительной системы и отвечает за восприятие светлого времени суток как времени бодрствования, а также поддерживает суточный ритм.
 Бодрствование - состояние психики, характеризующееся достаточно высоким уровнем электрической активности мозга, свойственным активному взаимодействию индивида с внешним миром. В поддержании состояния бодрствования важнейшую роль играет ретикулярная формация среднего мозга, от нейронов которой восходящие влияния идут к неспецифическим ядрам таламуса, а от них ко всем зонам коры больших полушарий. Бодрствование (обычное состояние не спящего человека) образует поле всевозможных сочетаний функций сознания - от состояния спокойного бодрствования через активное, напряженное бодрствование до выраженных аффектов.
Бодрствование - состояние психики, характеризующееся достаточно высоким уровнем электрической активности мозга, свойственным активному взаимодействию индивида с внешним миром. В поддержании состояния бодрствования важнейшую роль играет ретикулярная формация среднего мозга, от нейронов которой восходящие влияния идут к неспецифическим ядрам таламуса, а от них ко всем зонам коры больших полушарий. Бодрствование (обычное состояние не спящего человека) образует поле всевозможных сочетаний функций сознания - от состояния спокойного бодрствования через активное, напряженное бодрствование до выраженных аффектов.


 Сон – это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью.
Сон – это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью.
 Выделяют две основные стадии сна - «медленный» и «быстрый» сон. Феномен «быстрого» сна открыли в 1953 г. американские ученые А. Азеринский и К. Клейтман. Медленноволновой сон человека периодически прерывается короткими периодами низкоамплитудной ЭЭГ (похожей на десинхронизацию ЭЭГ при пробуждении), сопровождающимися быстрыми движениями глазных яблок. Отсюда часто употребляемое название этой стадии - парадоксальная, или стадия быстрых движений глаз (БДГ-сон, или REM-стадия - от rapid eye movement). Люди, разбуженные во время парадоксальной стадии сна, в 80 % случаев сообщали о сновидениях. Мышцы тела расслаблены сильнее всего во время парадоксального сна, однако вегетативные показатели (дыхание, сердцебиение, кровяное давление и др.) во время парадоксального сна могут соответствовать активному бодрствованию (вплоть до так называемой «вегетативной бури»).
Выделяют две основные стадии сна - «медленный» и «быстрый» сон. Феномен «быстрого» сна открыли в 1953 г. американские ученые А. Азеринский и К. Клейтман. Медленноволновой сон человека периодически прерывается короткими периодами низкоамплитудной ЭЭГ (похожей на десинхронизацию ЭЭГ при пробуждении), сопровождающимися быстрыми движениями глазных яблок. Отсюда часто употребляемое название этой стадии - парадоксальная, или стадия быстрых движений глаз (БДГ-сон, или REM-стадия - от rapid eye movement). Люди, разбуженные во время парадоксальной стадии сна, в 80 % случаев сообщали о сновидениях. Мышцы тела расслаблены сильнее всего во время парадоксального сна, однако вегетативные показатели (дыхание, сердцебиение, кровяное давление и др.) во время парадоксального сна могут соответствовать активному бодрствованию (вплоть до так называемой «вегетативной бури»).

 График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека. Видно резкое сокращение длительности парадоксального сна (БДГ) - от 8 часов у новорожденных до 1 часа у пожилых людей. Изменения в продолжительности медленноволнового сна (МВ) не столь выражено; уменьшение от 8 часов до примерно 5 часов.
График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека. Видно резкое сокращение длительности парадоксального сна (БДГ) - от 8 часов у новорожденных до 1 часа у пожилых людей. Изменения в продолжительности медленноволнового сна (МВ) не столь выражено; уменьшение от 8 часов до примерно 5 часов.
 ЭЭГ человека при бодрствовании и сне Бодрствование Частота Амплитуда Синхронизация (регулярность колебаний) Медленноволновый сон Парадоксальный сон (БДГ) высокая низкая (нерегулярная активность, десинхронизация) высокая (синхронизированная активность) низкая (нерегулярная активность, десинхронизация)
ЭЭГ человека при бодрствовании и сне Бодрствование Частота Амплитуда Синхронизация (регулярность колебаний) Медленноволновый сон Парадоксальный сон (БДГ) высокая низкая (нерегулярная активность, десинхронизация) высокая (синхронизированная активность) низкая (нерегулярная активность, десинхронизация)
 В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же ритм наблюдается при парадоксальном сне. В процессе засыпания человек проходит 4 стадии, от спокойного бодрствования с закрытыми глазами (альфа-ритм) до собственно медленноволнового сна (дельта-ритм).
В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же ритм наблюдается при парадоксальном сне. В процессе засыпания человек проходит 4 стадии, от спокойного бодрствования с закрытыми глазами (альфа-ритм) до собственно медленноволнового сна (дельта-ритм).
 Основные ритмы ЭЭГ человека Частота Состояние 12 -25 Гц Бодрствование с открытыми глазами и БДГ-сон Альфа-ритм 8 -12 Гц Бодрствование с закрытыми глазами Тета-ритм 4 -8 Гц Засыпание 1 -4 Гц Медленноволновый сон Бета-ритм Дельта-ритм
Основные ритмы ЭЭГ человека Частота Состояние 12 -25 Гц Бодрствование с открытыми глазами и БДГ-сон Альфа-ритм 8 -12 Гц Бодрствование с закрытыми глазами Тета-ритм 4 -8 Гц Засыпание 1 -4 Гц Медленноволновый сон Бета-ритм Дельта-ритм
 ЭЭГ при различных уровнях бодрствования и сна. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды не регистрируют мышечной активности нигде, кроме глазных мышц.
ЭЭГ при различных уровнях бодрствования и сна. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды не регистрируют мышечной активности нигде, кроме глазных мышц.
 Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими записями ЭЭГ (так называемые «перерезки Бремера» - по Бремеру, 1937). При перерезке на границе головного мозга со спинным (препарат А) в ЭЭГ сохранялась картина бодрствования, а при перерезке на уровне среднего мозга (препарат Б) получали препарат спящего мозга. Бремер заключил, что в изолированном головном мозгу имеется центр пробуждения, расположенный в продолговатом и среднем мозгу.
Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими записями ЭЭГ (так называемые «перерезки Бремера» - по Бремеру, 1937). При перерезке на границе головного мозга со спинным (препарат А) в ЭЭГ сохранялась картина бодрствования, а при перерезке на уровне среднего мозга (препарат Б) получали препарат спящего мозга. Бремер заключил, что в изолированном головном мозгу имеется центр пробуждения, расположенный в продолговатом и среднем мозгу.
 Дж. Моруцци и Х. Мегун показали в 1949 г. , что стимуляция ретикулярной формации ствола мозга вызывает пробуждение. Сон при этом в то время рассматривался как следствие временной блокады активирующих восходящих влияний с одновременным «включением» таламокортикальных синхронизирующих процессов.
Дж. Моруцци и Х. Мегун показали в 1949 г. , что стимуляция ретикулярной формации ствола мозга вызывает пробуждение. Сон при этом в то время рассматривался как следствие временной блокады активирующих восходящих влияний с одновременным «включением» таламокортикальных синхронизирующих процессов.
 В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации» (представление о которой сформировалось благодаря работам Бремера, Моруцци, Мегуна и других исследователей), существует большое количество центров, которые различаются по своим функциям, выделяемым нейромедиаторам и локализации в мозге. Большинство из них действительно находятся в ретикулярной формации ствола мозга или вблизи нее, но часть из них – в гипоталамусе и других структурах мозга.
В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации» (представление о которой сформировалось благодаря работам Бремера, Моруцци, Мегуна и других исследователей), существует большое количество центров, которые различаются по своим функциям, выделяемым нейромедиаторам и локализации в мозге. Большинство из них действительно находятся в ретикулярной формации ствола мозга или вблизи нее, но часть из них – в гипоталамусе и других структурах мозга.
 Некоторые системы регуляции сна и бодрствования. Видно, как в цепях, выходящих из голубого пятна (медиатор - норадреналин) и ядер шва (медиатор - серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
Некоторые системы регуляции сна и бодрствования. Видно, как в цепях, выходящих из голубого пятна (медиатор - норадреналин) и ядер шва (медиатор - серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
 Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и сна Бодрствование Медленноволновый сон Парадоксальный сон Преобладающий ритм в ЭЭГ бета-ритм дельта-ритм бета-ритм Выделение норадреналина (из голубого пятна) максимально снижено отсутствует Выделение серотонина (из дорзальных ядер шва) максимально снижено отсутствует Выделение гистамина (из туберомамиллярного ядра заднего гипоталамуса) максимально снижено отсутствует Выделение ацетилхолина (из базальных ядер переднего мозга - базальное крупноклеточное ядро и др.) максимально отсутствует максимально
Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и сна Бодрствование Медленноволновый сон Парадоксальный сон Преобладающий ритм в ЭЭГ бета-ритм дельта-ритм бета-ритм Выделение норадреналина (из голубого пятна) максимально снижено отсутствует Выделение серотонина (из дорзальных ядер шва) максимально снижено отсутствует Выделение гистамина (из туберомамиллярного ядра заднего гипоталамуса) максимально снижено отсутствует Выделение ацетилхолина (из базальных ядер переднего мозга - базальное крупноклеточное ядро и др.) максимально отсутствует максимально
 В настоящее время очевидно, что бодрствование и сон – качественно различные состояния, каждое из которых имеет свои управляющие центры.
В настоящее время очевидно, что бодрствование и сон – качественно различные состояния, каждое из которых имеет свои управляющие центры.
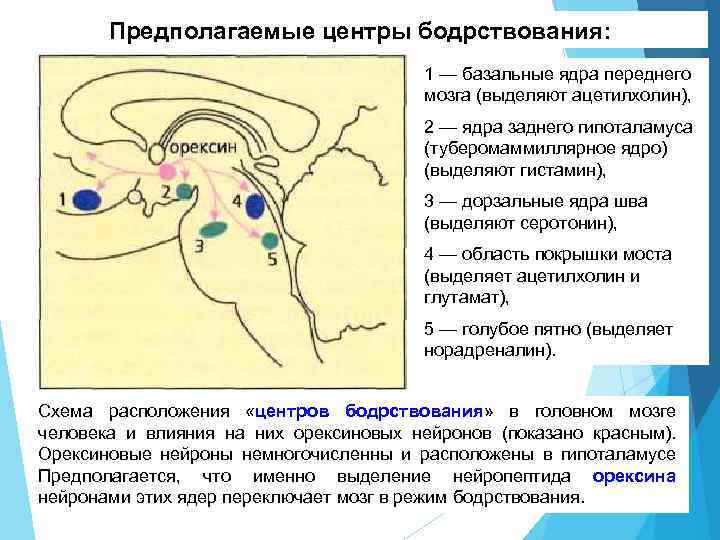 Предполагаемые центры бодрствования: 1 - базальные ядра переднего мозга (выделяют ацетилхолин), 2 - ядра заднего гипоталамуса (туберомаммиллярное ядро) (выделяют гистамин), 3 - дорзальные ядра шва (выделяют серотонин), 4 - область покрышки моста (выделяет ацетилхолин и глутамат), 5 - голубое пятно (выделяет норадреналин). Схема расположения «центров бодрствования» в головном мозге человека и влияния на них орексиновых нейронов (показано красным). Орексиновые нейроны немногочисленны и расположены в гипоталамусе Предполагается, что именно выделение нейропептида орексина нейронами этих ядер переключает мозг в режим бодрствования.
Предполагаемые центры бодрствования: 1 - базальные ядра переднего мозга (выделяют ацетилхолин), 2 - ядра заднего гипоталамуса (туберомаммиллярное ядро) (выделяют гистамин), 3 - дорзальные ядра шва (выделяют серотонин), 4 - область покрышки моста (выделяет ацетилхолин и глутамат), 5 - голубое пятно (выделяет норадреналин). Схема расположения «центров бодрствования» в головном мозге человека и влияния на них орексиновых нейронов (показано красным). Орексиновые нейроны немногочисленны и расположены в гипоталамусе Предполагается, что именно выделение нейропептида орексина нейронами этих ядер переключает мозг в режим бодрствования.
 Функция орексина как медиатора, ответственного за координацию других центров бодрствования, была открыта совсем недавно – около 2003 г. , а сам орексин был открыт лишь в 1998 г. При потере хотя бы части орексиновых нейронов возникает нарколепсия – дневная сонливость.
Функция орексина как медиатора, ответственного за координацию других центров бодрствования, была открыта совсем недавно – около 2003 г. , а сам орексин был открыт лишь в 1998 г. При потере хотя бы части орексиновых нейронов возникает нарколепсия – дневная сонливость.
 Схема центра расположения центра медленного сна. Вверху – бодрствование, когда центр сна заторможен и ретикулярная формация (РФ) активирует кору; внизу – центр сна возбужден, РФ заторможена и кора не активируется.
Схема центра расположения центра медленного сна. Вверху – бодрствование, когда центр сна заторможен и ретикулярная формация (РФ) активирует кору; внизу – центр сна возбужден, РФ заторможена и кора не активируется.
 Предполагаемые центры сна: Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом ядре), медиатор - гаммааминомасляная кислота (ГАМК); активность нейронов незначительна в бодрствовании, но резко возрастает в период обычного сна и прекращается во время парадоксального. Центр парадоксального сна: ретикулярная формация покрышки моста (латеродорзальная / педункуло-понтийная области покрышки моста), медиаторы - ацетилхолин и глутамат.
Предполагаемые центры сна: Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом ядре), медиатор - гаммааминомасляная кислота (ГАМК); активность нейронов незначительна в бодрствовании, но резко возрастает в период обычного сна и прекращается во время парадоксального. Центр парадоксального сна: ретикулярная формация покрышки моста (латеродорзальная / педункуло-понтийная области покрышки моста), медиаторы - ацетилхолин и глутамат.
 Список Литературы Дубынин В. А. Регуляторные системы организма человека. М. : Дрофа, 2003. Дудьев В. П. Психомоторика: cловарь-справочник, 2008 г. Мэгун Г. Бодрствующий мозг. М. : Мир, 1965. Росси Я. А. , Цанкетти А. Ретикулярная формация ствола мозга. М. : ИЛ, 1960. Шульговский В. В. Физиология центральной нервной системы. М. : Изд-во Моск. ун-та, 1987.
Список Литературы Дубынин В. А. Регуляторные системы организма человека. М. : Дрофа, 2003. Дудьев В. П. Психомоторика: cловарь-справочник, 2008 г. Мэгун Г. Бодрствующий мозг. М. : Мир, 1965. Росси Я. А. , Цанкетти А. Ретикулярная формация ствола мозга. М. : ИЛ, 1960. Шульговский В. В. Физиология центральной нервной системы. М. : Изд-во Моск. ун-та, 1987.
Переход от бодрствования ко сну предполагает два возможных пути. Прежде всего не исключено, что механизмы, поддерживающие бодрствующее состояние, постепенно «утомляются». В соответствии с такой точкой зрения сон - это пассивное явление, следствие снижения уровня бодрствования. Однако не исключено и активное торможение обеспечивающих бодрствование механизмов. Важную роль в регуляции цикла сон - бодрствование играет ретикулярная формация ствола мозга, где находится множество диффузно расположенных нейронов, аксоны которых идут почти ко всем областям головного мозга, за исключением новой коры (неокортекса). Ее роль в цикле сон - бодрствование была исследована в конце 40-х годов Г. Моруцци и Н. Мэгуном. Они обнаружили, что высокочастотное электрическое раздражение этой структуры у спящих кошек приводит к их мгновенному пробуждению. И напротив, повреждения ретикулярной формации вызывают постоянный сон, напоминающий кому; перерезка же только сенсорных трактов, проходящих через ствол мозга, такого эффекта не дает.
Серотонинергические нейроны также играют весьма важную роль в регуляции бодрствования и сна. В верхних отделах ствола мозга есть две области - ядра шва и голубое пятно , для нейронов которых характерны такие же обширные проекции, как и для нейронов ретикулярной формации, т.е. достигающие многих областей ЦНС. Медиатором в клетках ядер шва служит серотонин (5-гидрокситриптамин, 5-НТ ), а голубого пятна - норадреналин . Разрушение ядер шва у кошки приводит к полной бессоннице в течение нескольких дней; за несколько следующих недель сон нормализуется. Частичная бессонница может быть также вызвана подавлением синтеза 5-НТ n -хлорфенилаланином. Ее можно устранить введением 5-гидрокситриптофана, предшественника серотонина, не проникающего через гематоэнцефалический барьер. Двустороннее разрушение голубого пятна приводит к полному исчезновению БДГ-фаз, не влияя на медленноволновой сон. Истощение запасов серотонина и норадреналина под влиянием резерпина вызывает, как и следовало ожидать, бессонницу. Однако оказалось, что нейроны ядер шва наиболее активны и выделяют максимум серотонина не во время сна, а при бодрствовании . Кроме того, возникновение БДГ, по-видимому, обусловлено активностью нейронов не столько голубого пятна, сколько более диффузного подголубого ядра. Судя по результатам недавних экспериментов, серотонин служит и медиатором в процессе пробуждения, и «гормоном сна» в бодрствующем состоянии, стимулируя синтез или высвобождение «веществ сна» , (факторов сна), которые в свою очередь вызывают сон. Были сделаны попытки обнаружить особые вещества либо после длительного лишения сна, либо у спящего человека. Первая из этих попыток основана на предположении о том, что фактор(ы) сна во время бодрствования накапливаются до вызывающего сон уровня, а вторая - на гипотезе, согласно которой они образуются или выделяются во сне. Исследования дали определенные результаты. Так, при проверке первой гипотезы из мочи и цереброспинальной жидкости человека и животных был выделен небольшой глюкопептид -фактор S, вызывающий медленноволновой сон при введении другим животным. Существует, по-видимому, и фактор сна с БДГ. Изучение второй гипотезы привело к открытию индуцирующего глубокий сон нонапептида (в настоящее время он уже синтезирован), так называемого пептида д-сна (DSIP , delta-sleep inducing peptide). Однако пока не известно, играют ли эти и многие другие «вещества сна», обнаруженные при проверке обеих гипотез, какую-либо роль в его физиологической регуляции. Более того, выделенные пептиды часто вызывают сон лишь у животных определенного вида; кроме того, он возникает и под действием других веществ. Однако сросшиеся девочки-близнецы могли спать порознь, что свидетельствует о второстепенной роли гуморальных факторов и решающей роли в развитии сна нервной системы.



